
Les insectes sont les premiers organismes à avoir développé la capacité de voler, il y a 170 millions d’années (lire article sur la phylogénie). L’acquisition du vol constitua une innovation majeure dans leur évolution : elle leur a procuré un avantage décisif comme échapper à la prédation, fuir un environnement hostile pour en rechercher un plus favorable et aussi rencontrer des individus du sexe opposé pour la reproduction.
Contrairement à l’aile des vertébrés, celle des insectes ne possède pas de muscles intrinsèques (à l’intérieur de l’organe), elle est rattachée au thorax par un ensemble complexe d’éléments articulés appelés pteralia (lire cet article sur l’anatomie de l’aile).
Leonard de Vinci a déterminé que le vol nécessitait trois éléments : 1) Un moteur léger et puissant ; 2) Des ailes capables de générer des forces aérodynamiques suffisantes ; 3) Un système de contrôle perfectionné pour maintenir le corps en l’air. Les insectes satisfont ces trois critères.
Malgré d’importantes recherches sur le vol des insectes, celui-ci demeure encore peu expliqué car difficile à observer en raison de la taille de ces animaux et de la vitesse des mouvements. Il reste un sujet complexe et vaste. Avec cet article, j’ai voulu synthétiser trois points essentiels pour vous permettre d’en apprendre davantage sur : l’aérodynamisme, la musculature et le système de contrôle.
I) Ailes et aérodynamisme
Le vol des insectes est bien plus compliqué que celui des avions qui repose sur 2 principes :
- Principe de Bernouilli : l’aile est profilée de telle manière qu’en se déplaçant, elle provoque une division du flux d’air : le courant circulant au dessus, en accélérant, crée une dépression et donc une aspiration (force perpendiculaire à la surface de l’aile), ce phénomène est appelé la portance.
- La Portance : l’inclinaison de l’aile (son angle d’attaque) augmente la force d’aspiration, au delà de 15°, cette portance disparaît, entrainant le décrochage de l’avion et donc sa chute.
Les principes physiques conditionnant le vol repose sur la mécanique des fluides car un gaz se comporte comme un liquide visqueux.
Chez les insectes, l’aile est plane et présente un angle d’attaque de 30-40°, donc bien supérieur au 15° : Comment peuvent-ils se maintenir en vol dans ces conditions?
I) 1 : Le battement des ailes
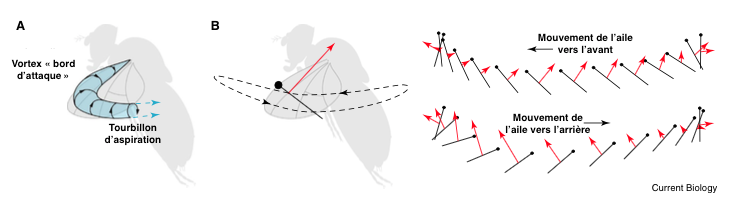
Contrairement aux idées reçues, les insectes, hormis les libellules, ne battent pas des ailes de haut en bas, mais pratiquement sur un plan horizontal d’avant en arrière (mouvement sinusoïdal). Lorsque l’insecte bat de l’aile, il se forme un « vortex bord d’attaque » : un tourbillon de l’air, au niveau du bord d’attaque de l’aile ce qui engendre un phénomène d’aspiration perpendiculaire à la surface alaire (voir figure 1). L’angle d’attaque étant supérieur au 15°, l’insecte décroche aussitôt, amorce une chute qui sera très rapidement compensée par un nouveau battement.
L’insecte se maintient donc en vol grâce à des battements d’ailes à très haute fréquence mais ce seul phénomène ne suffit pas à expliquer son vol.
Les insectes pourraient diminuer leur angle d’attaque et augmenter l’efficacité en terme d’aspiration, mais au prix d’une fréquence de battement supérieure, au delà des capacités physiques et biologiques de la contraction musculaire (voir chapitre suivant). Aussi, les insectes doivent accepter un compromis et sont obligés de composer avec les contraintes physiques et biologiques.
A l’échelle à laquelle évolue les insectes, la viscosité de l’air entre également en compte.
I) 2 : La viscosité de l’air
La performance aérodynamique de l’aile et la portance sont liées à la façon dont s’écoule l’air, donc le fluide, suite au passage de l’aile. Ce phénomène est caractérisé par le nombre de Reynolds : Re (voir encadré). Ainsi, lorsque Re est inférieur à 2000, les forces liées à la viscosité de l’air dominent, l’écoulement est laminaire (sans turbulence), au dessus de 3000 les forces liées à la portance dominent et l’écoulement devient turbulent (avec des perturbations).
| Le nombre de Reynolds (de l’ingénieur irlandais Osborne Reynolds, 1883) est en sans dimension, il permet de déterminer le régime d’écoulement d’un fluide, qui peut être : laminaire, transitoire ou turbulent. Il donne une mesure du rapport des forces d’inertie aux forces visqueuses, par conséquent permet de quantifier l’importance relative des ces deux types de forces pour des conditions déroulement données. |
Compte tenu de leur taille, des vitesses atteintes et de la viscosité de l’air, les ailes d’insectes ont un chiffres de Reynolds très faible, comprises entre 10 et 5000, pour comparaison, le Reynolds d’une aile d’avion se situe à plusieurs millions. Ces paramètres procurent un avantage à l’insecte en lui offrant une portance plus élevée que pour des Reynolds plus grands.
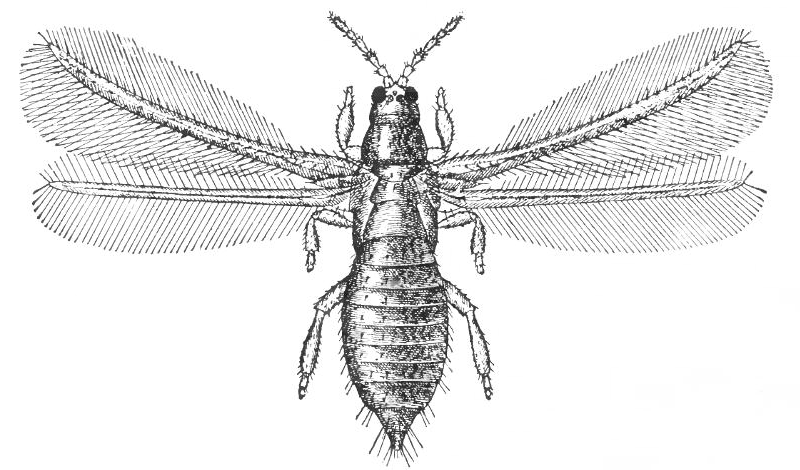
L’évolution a engendré la miniaturisation, moins de 1mm pour certaines espèces comme les Thrips (Thysanoptères) ou les guêpes parasitoïdes (Hyménoptères). A cette échelle, les lois physiques sont encore différentes car le Reynolds est encore plus faible et la viscosité particulièrement forte. Ces insectes possèdent des adaptations particulières sur les ailes, elles sont en forme de « brosse » avec une frange ciliée dont le rôle n’est pas encore élucidé. Ces structures auraient pour fonction de générer un excès de trainée visqueuse (voir illustration 1 ci-contre).
I) 3 : Capture de sillage
Les insectes font appel à un autre phénomène : le « capture de sillage » (« Wake Capture« ).
En se déplaçant, l’aile provoque dans son sillage des perturbations et des turbulences de l’air. Lors de son mouvement retour, l’insecte profite d’une partie de cette énergie et des forces générées pour augmenter sa portance et son aspiration (voir figure 2).
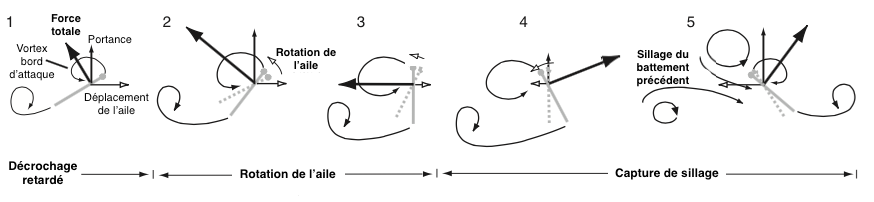
L’utilisation de ce phénomène confère un avantage à l’insecte malgré le fait que les turbulences et les perturbations compliquent le contrôle, la sensibilité et la maniabilité du vol.
Bien que certains principes de l’aérodynamique s’appliquent à l’ensemble des insectes, les particularités et les adaptations vont de pair avec la diversité morphologique et comportementale des espèces. De plus, l’aile n’a pas pour seul but de faire voler l’insecte, elle doit également lui permettre de maintenir une direction et une vitesse déterminées tout en compensant les turbulences et les dérives dues aux courants aériens.
L’étude de l’aérodynamique du vol des insectes doit être mis en relation avec leur aptitude à la manoeuvre et leur comportement pour la survie et la reproduction.
II) Les muscles
La capacité de voler a été rendue possible grâce à l’apparition d’une musculature à la fois puissantes et pouvant se contracter à de très grandes fréquences. Au cours de l’évolution, il y a eu miniaturisation chez beaucoup de familles comme les diptères, les hymènoptères, les coléoptères, etc. En raison des principes physiques, plus les espèces sont petites, plus elles doivent battre rapidement des ailes pour combattre la gravité. A cette échelle, la viscosité de l’air augmente et la performance aérodynamique faiblit. De nombreuses adaptations anatomiques et physiologiques sont permis de pallier à ces contraintes.
Exemple : Les ailes des grands papillons comme les sphinx (Sphingidae) battent à une fréquence de 30Hz et les moustiques à plus de 1000Hz.
Or, la limite biologique de la fréquence des contractions musculaires se situe en deçà de ces valeurs. Comment les insectes ont-ils pu contourner cette problématique?
Pour rappel, l’arrivée d’un influx nerveux acheminé par un motoneurone, à une cellule musculaire provoque une succession d’événements : 1) Libération d’ions calcium Ca2+ du réticulum sarcoplasmique (RS) ; 2) Liaison des ions Ca2+ à une protéine : la troponine ; 3) Celle-ci engendre le déplacement de la tropomyosine ; 4) La tropomyosine libère à son tour la myosine qui peut se lier à l’actine, déclenchant finalement la contraction des fibres contractiles de la cellule musculaire (voir vidéo en bas de page).
Cette chaine chimique est particulièrement rapide car le calcium possède une force électrochimique très forte (il diffuse passivement dans le sens du gradient de concentration ou gradient électrochimique). Le retour à la situation de départ, la désactivation de la cellule musculaire, est quant à lui plutôt lent car il fait appel à un afflux actif des Ca2+ dans les RS (sens opposé au gradient) : ce processus limite la fréquence de contraction des muscles alaires.
Les muscles des vertébrés et de certaines espèces d’insectes possèdent des cellules musculaires à forte concentration en RS pour offrir de la rapidité, au détriment des fibres contractiles et des mitochondries responsables de la puissance. Il existe donc un compromis puissance/rapidité lié aux contraintes physiques et physiologiques.
Ainsi, durant l’histoire évolutive des insectes, des innovations ont vu le jour pour répondre à ces contraintes. Une des principales d’entres elles est l’apparition de nouveaux types de muscles, appelés « asynchrones » ou moteurs. Cette innovation est survenue à quatre reprises au sein des différentes lignées.
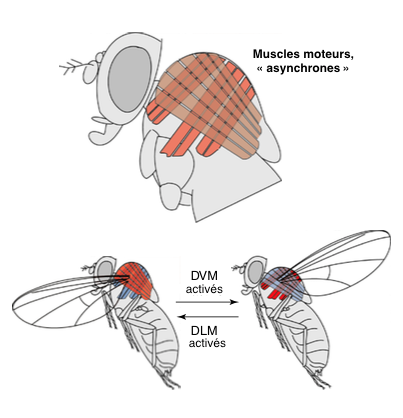
Le principe d’asynchronie consiste à faire fonctionner deux groupes de muscles antagonistes successivement :
- muscles dorso-longitudinaux (MDL)
- muscles dorso-ventraux (MDV)
Ces muscles, « indirects », ne sont pas liés aux ailes mais s’insèrent à la parois interne des éléments constituant l’exosquelette du thorax (lire cet article).
La contraction des MDL déplacent l’aile vers l’avant, ce qui active les MDV et enclenche le mouvement retour (voir figures 3 et 4). Ce mécanisme permet de ne solliciter qu’un des deux groupes de muscles à la fois, et permet de doubler la fréquence de battement.
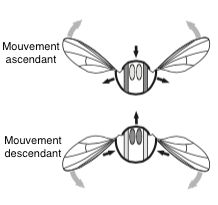
Ces muscles « indirects » vont fournir la puissance et l’énergie mécanique suffisante pour permettre à l’insecte de voler.
Le contrôle de la trajectoire va être réalisé par douze muscles, dits « directs » car reliés à la base de l’aile. Ces muscles, contrairement aux muscles « indirects », sont synchrones et ne sont pas capables de générer de grandes forces et des vitesses de contraction rapide en raison d’une concentration de réticulum sarcoplasmique dans leurs cellules.
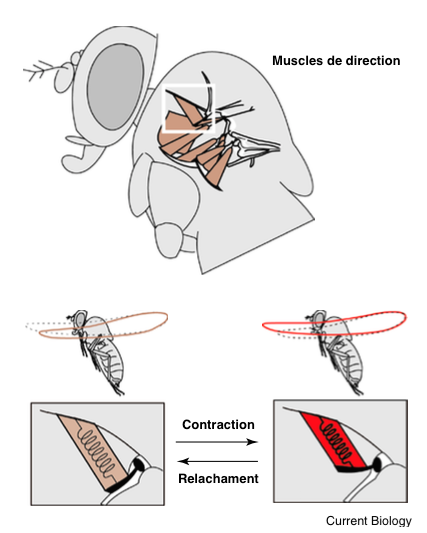
Par contre, ils agissent de manière très fine sur l’aile pour modifier la forme de sa base, la cinétique et la cinématique de sont mouvement, ce qui va permettre à l’insecte de changer de trajectoire de vol. Ils jouent en quelque sorte le rôle des ressorts des volets situés à l’arrière des ailes d’avion (voir figure 5).
Le vol impose l’analyse de nombreux paramètres (vitesse de l’animal et des courants aériens, position tridimensionnelle dans l’espace, altitude, informations visuelles…). L’évolution a fourni aux insectes un système nerveux à la fois rapide et performant.
III) Le système de contrôle
Les insectes volant diurnes comme les libellules, les mouches, les guêpes et les papillons ont un système nerveux fortement spécialisés : des yeux composés et des ganglions visuels hypertrophiés dont le rôle est d’informer l’insecte sur la vitesse et l’altitude, de permettre l’atterrissage et d’éviter des obstacles ou des prédateurs, ainsi qu’une bonne mémoire spatiale.
Le système d’analyse des insectes est l’un des plus rapides et des plus efficaces du monde animale. Le traitement de l’information visuelle fonctionne à une fréquence de 300Hz.
Le système de contrôle de vol est composé de :
- d’ocelles (oeil simple sensible aux variations de luminosité) présent sur le sommet de la tête dont le rôle est de détecter tout changement dans la position horizontale du corps et donc de contenir le tangage et le roulis
- de soies et de mécanorécepteurs présents sur la tête dont le rôle est de mesurer l’amplitude et la vitesse de l’air durant le vol
- de nombreux organes à la base de l’aile comme des organes campaniformes (détection de la déformation de la structure de la cuticule), des soies et des tegulae (petites « écailles » présentes à la surface de l’aile)
- des haltères chez les diptères dont le rôle est de contrer la force de coriolis (lire cet article et voir vidéo en fin d’article).
Des articles décrivant l’ensemble des organes sensoriels de l’insecte seront publiés prochainement. Celui sur les mécanorécepteurs sensoriels cuticulaire est à lire ici.

Un problème biologique demeure cependant pour les petites espèces. La fréquence à laquelle les ailes battent est bien trop rapide pour être contrôlée par le système nerveux central (cerveau). La vitesse de transmission nerveuse de l’information et de son analyse entre les ailes et le cerveau requiert trop de temps en raison des nombreuses chaines de réactions chimiques successives nécessaires. Pour remédier à ce problème, l’évolution a développé un système autonome dans le thorax basé sur des boucles réflexes (voir figure 6).
La contraction des muscles de direction, situés à la base de l’aile, sont directement contrôlés par des motoneurones provenant des haltères (ailes postérieures modifiés chez les diptères) et des autres organes sensorielles de l’aile et de sa base renseignant sur les paramètres de vol. Ainsi, la position, la cinétique et la cinématique de l’aile sont modifiées sans que le traitement de l’information ait à se déplacer jusqu’au cerveau. Ce procédé permet de gagner en rapidité, donc en performance et en efficacité.
L’intégration de ces deux systèmes nerveux (système central et des ailes) n’est toutefois pas encore bien comprise. De nombreuses zones d’ombres demeurent.
Une étude scientifique récente (2015) montre que chez les espèces n’ayant pas d’haltères, les ailes interviennent dans la détection et le contrôle de la force de coriolis, elles jouent ainsi le rôle de gyroscopes. Pour en savoir plus, lire cet article.
Les études sur le vol des insectes s’inscrivent dans un objectif de concevoir des microdrones, inférieurs à 15cm, pour des besoins civils et militaires. Ces machines doivent avoir la capacité d’effectuer du vol stationnaire et offrir une bonne manoeuvrabilité.
Le vol battu des insectes semble être la solution la plus adaptée. Les ingénieurs travaillent sur le biomimétisme, qui consiste à s’inspirer des modèles biologiques pour la conception de systèmes mécaniques.
Vidéo : Explication du vol des insectes par Mickael Dickinson :
Vidéo : Fonctionnement des haltères chez les diptères :
Vidéo : La contraction musculaire :
Sources :
- Michael Dickinson (2011) : Insect flight. Current Biology, vol 16 N°9 (lien)
- Sane S.P. (2003) : The aerodynamics of insect flight. The Journal of Experimental Biology, 206 : 4191-4208 (lien)
- Evolution of the Insects ; D. GRIMALDI & M.S. ENGEL, Cambridge University Press-2005
- Encyclopedia of Insects ; Vincent H. RESH & Ring T. CARDE, Academic Press-2009
Recommandation d’ouvrages sur cette thématique
– Evolution of the Insects (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
– The Insects: An Outline of Entomology (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
– The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
– Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
<img src= »http://wms-eu.amazon-adsystem.com/20070822/FR/img/noscript.gif?tag=passioentomo-21&linkId=3LHYKSRXQKZ3KY5I » alt= » » /><br />
Bonsoir,
je ne suis pas scientifique mais je cherche à comprendre la trajectoire du papillon. Nous entendons souvent dire d’une personne dispersée qu’elle papillonne. Est-ce qu effectivement le papillon ne vont pas d’un point A a un point B en y allant directement sur une ligne droite. Ou alors ont-ils un plan de vol un peu étrange pour y arriver parce qu’ils ne peuvent pas faire autrement ? J’aimerai trouver des schémas techniques expliquant cette fameuse trajectoire dispersée ou pas (si il y a une logique).
merci pour votre réponse
Audrey