Première partie : les mécanorécepteurs cuticulaires
Comme l’ensemble des arthropodes, les insectes ont un squelette externe, l’exosquelette, constitué de chitine et formant la cuticule (lire cet article). Ses propriétés isolantes impliquent que la cuticule crée une barrière à la réception des stimuli de l’environnement extérieur vers le milieu physiologique interne. Afin de contourner cette problématique, l’évolution a fait surgir tout un ensemble de structures destinées à transmettre une large gamme de types de stimuli à l’insecte qui lui permettent d’appréhender de manière efficace et précise son environnement et de répondre favorablement à toute modification par un comportement adapté.
Parmi ces structures sensorielles, certaines sont sensibles aux distorsions mécaniques (mécanorécepteurs : objet de cette article), d’autres à la lumière (photorécepteurs) et d’autres encore à des composants chimiques (chémorécepteurs ou chimiorécepteurs).
I) Généralités
Les mécanorécepteurs sont classés en trois grandes catégories (Tableau I) :
| Catégories | Position anatomique | Types | Fonctions |
| Extérorécepteurs Récepteurs cuticulaires | Surface de la cuticule | Sensilles simples | Toucher, variation de pression (air ou eau), détection de la gravité, proprioception* |
| Sensilles campaniformes | Variation de pression sur la cuticule | ||
| Intérorécepteurs Récepteurs intradermiques ou subcuticulaires (cliquez ici) | Derme | Organes chordotonaux | Variation de pression (air, eau), détection de la gravité, proprioception* |
| Propriorécepteurs | Tissus internes | Pas de structure précise | Proprioception* |
| * Proprioception : perception de la position des différentes parties du corps | |||
Dotés de propriétés étonnantes et multiples, ils peuvent intervenir dans le sens du toucher lorsqu’ils sont stimulés par le contact d’un objet ou un substrat, dans le sens de l’ouïe par une stimulation de l’air ou de l’eau et aussi dans le contrôle de la posture et des mouvements quand ils sont excités par des déplacements de parties du corps de l’insecte.
Deux types de mécanorécepteurs cuticulaires se rencontrent chez les insectes : les sensilles simples (poils à la surface de la cuticule) et les sensilles campaniformes, structure en forme de dôme où la cuticule est de faible épaisseur.
Malgré une structure différente, l’organisation cellulaire des deux types de sensilles est similaire (voir paragraphe correspondant).
II) Description structurelle
II) 1. Sensilles simples
La plupart des poils qui recouvrent le corps des insectes sont des mécanorécepteurs isolés ou regroupés. Selon les espèces et leur fonction, ces sensilles, de longueur variables, peuvent se terminer en pointe, ou non (et sont alors appelées les trichobotries). L’extrémité d’autres sensilles se terminent en forme de petite massue : elles interviennent dans la détection du champ magnétique.
L’évolution a permis l’apparition d’autant de variantes qu’il y a de fonctions.
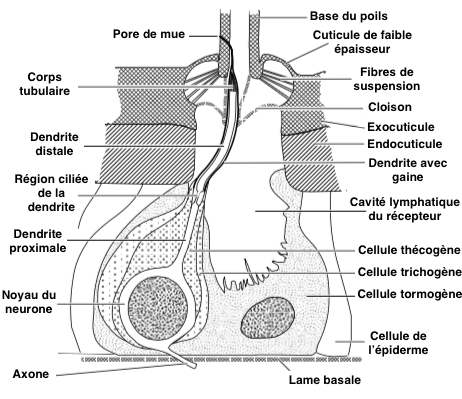
La structure anatomique des sensilles simples est identique quel qu’en soit le type (voir figure 1). La sensille est une excroissance de l’exocuticule dont la base est composée de chitine de faible épaisseur garantissant ainsi une certaine souplesse et le mouvement du poil. Des fibres élastiques de suspension placées à cette base permettent un retour à la position initiale après un déplacement. La sensille repose sur une cloison qui la sépare d’une cavité : la cavité lymphatique du récepteur.
Cette configuration est parfois spécialisée pour permettre à la sensille de détecter un déplacement dans une direction donnée : il s’agit alors d’une sensibilité directionnelle.
II) 2. Sensilles campaniformes
Ces structures ovalaires, d’un diamètre compris entre 5 et 30µm, sont constituées de deux couches cuticulaires : une couche externe mince et une couche interne fibreuse et souple. Parfois, une couche supplémentaire spongieuse s’insère entre ces couches (voir figure 2).
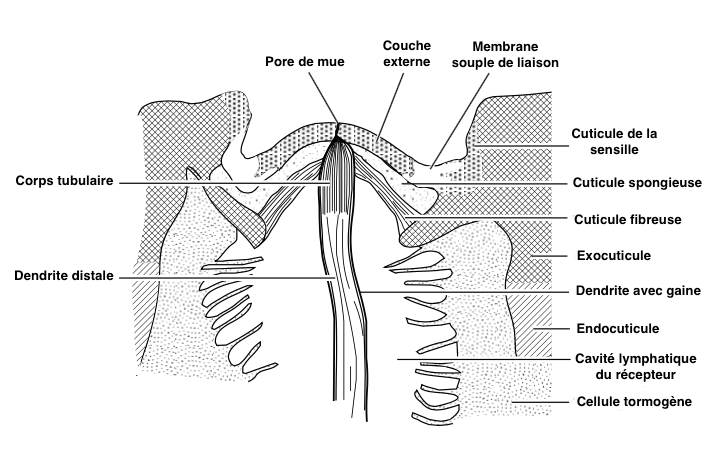
Les sensilles campaniformes sont réparties sur toute la surface du corps de l’insecte, notamment au niveau des articulations et des zones soumis à un stress mécanique important comme les tibias, les pièces buccales et les haltères des diptères (lire cet article et celui-ci).
Plusieurs variantes existent au sein d’un même individu selon la fonction. Ainsi, chez les mouches du genre Calliphora, sont présentes près de 1200 sensilles campaniformes réparties en neufs types : 36 par pattes, 140 à la base de chaque aile et 340 sur chaque haltère.
III) Description cellulaire
Les sensilles simples et campaniformes sont reliées à un complexe de plusieurs cellules épidermiques composé d’une cellule nerveuse (neurone) sensorielle et de deux ou trois cellules de soutien.
- Le neurone sensoriel, repérable à son noyau particulièrement volumineux, est rattaché à la base du poil de la sensille simple et inséré dans les deux couches cuticulaires de la sensille campaniforme par une seule dendrite non ramifiée et entourée d’une gaine (voir figure 1 & 2).
- Les cellules de soutien sont le plus souvent au nombre de trois : la thécogène (la plus proche du neurone) sécrète la gaine de la dendrite, la trichogène sécrète la cuticule du poil et la tormogène (la plus externe) sécrète la cloison de la cavité lymphatique du récepteur (voir figure 1). Chez certaines espèces, cette dernière peut disparaître une fois son rôle échu.
La dendrite est constituée de trois parties : une partie distale contenant de nombreuses mitochondries et autres organites cellulaires, une zone de rétrécissement ciliée au niveau de la cuticule (endocuticule) et, dans sa partie proximale, un corps tubulaire comprenant un grand nombre de microtubules (polymère de tubuline) qui le rendent extrêmement sensible aux variations de forme (voir figure 2).
IV) Fonctionnement
Le fonctionnement des mécanorécepteurs cuticulaires implique trois processus successifs : 1) le couplage ; 2) le potentiel de récepteur/générateur et 3) le potentiel d’action.
IV) 1. Le couplage
Le couplage est le mécanisme qui permet de recueillir l’énergie mécanique captée par le poil ou la sensille campaniforme.
Lorsque le poil de la sensille entre en contact avec un objet ou substrat, une force de flexion s’exerce à sa base, tel un levier : elle engendre une amplification du mouvement et entraine une distorsion mécanique au niveau du corps tubulaire du neurone sensoriel.
Le même principe s’observe dans la sensille campaniforme où le couplage est effectué par la compression ou la décompression de la cuticule à cet endroit. Certains corps tubulaires ont une forme d’éventail ce qui confère une sensibilité directionnelle à la sensille campaniforme.
Plusieurs types de sensilles sont présents sur le corps de l’insecte : il a été recensé jusqu’à 12 types sur une patte de blattes. Leurs variations en forme, en taille et en orientation, livrent à l’insecte une grande diversité d’informations.
VI) 2. Transduction & potentiel générateur
Ce processus correspond à la traduction de l’énergie mécanique en énergie électrique au niveau du corps tubulaire de la cellule nerveuse sensorielle. L’énergie électrique ainsi traduite est le potentiel générateur (PG), aussi appelé potentiel de récepteur.
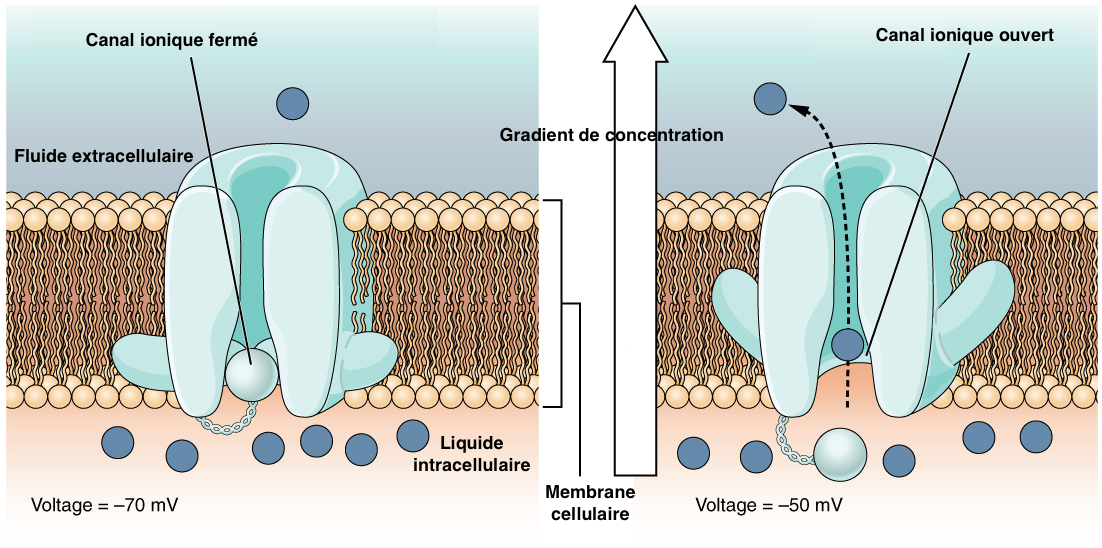
Une action mécanique sur le poil ou la sensille campaniforme provoque une déformation du corps tubulaire, entrainant l’ouverture de canaux ioniques (canaux à potassium le plus souvent) à la surface de la membrane plasmique. Cet échange d’ions crée une dépolarisation (passage transitoire d’une valeur négative à une valeur positive à la surface d’une cellule) et donc un courant électrique (voir figure 3).
La sensibilité est telle qu’une distorsion du corps de tubulaire d’à peine 5nm engendre un PG!
IV) 3. Potentiel d’action
Le potentiel d’action (PA) fait suite à une dépolarisation de la membrane plasmique au-delà d’une certaine valeur seuil. Aussi appelé influx nerveux, il correspond à la transmission de cette dépolarisation le long de la dendrite et du neurone, et donc de l’envoi du message nerveux à d’autres neurones à travers l’organisme.
Le processus consiste en une ouverture/fermeture successive des canaux ioniques modifiant la charge électrique de la membrane (voir vidéos ci-dessous). Ainsi, le potentiel membranaire passe d’une valeur de -70mV à +30mV puis retrouve sa valeur initiale pour être à nouveau activé. Ce cycle se déroule en moyenne en 2 millisecondes.
Chaque type de mécanorécepteurs possède un seuil de dépolarisation et de PA spécifique. La génération d’un PA dépend de la force s’exerçant sur le mécanorécepteur, de la sensibilité de couplage, de la nature des canaux ioniques, de l’intensité et de la fréquence de la dépolarisation.
La sensibilité et la réaction faisant suite à une stimulation du mécanorécepteur sont donc très variables.
Par ailleurs, la réponse à un stimulus peut-être de deux types : phasique ou tonique.
Une pression mécanique suffisante sur le mécanorécepteur entraine l’apparition d’un potentiel d’action (PA). Une réponse est dite phasique si le maintien de cette pression ne provoque pas de nouveau PA. Ce type de réponse concerne notamment les poils des sensilles simples.
Au contraire, s’il y a émission de plusieurs PA durant toute la phase de pression mécanique, la réponse sera tonique et typique de la réponse des sensilles campaniformes. Certaines sensilles campaniformes peuvent déclencher des réponses soit tonique soit phasique selon le type, la fréquence et l’intensité du stimulus.
Le traitement et l’activation des potentiels d’action sont régulés par de nombreux processus biochimiques et électriques. La réponse de l’insecte à toute une gamme de stimuli résulte de l’analyse de l’ensemble des stimuli, de leur intensité, de leur fréquence et de leur amplitude à tout instant.
V) Exemples du rôle joué par les mécanorécepteurs cuticulaires
Les poils de sensilles simples jouent un rôle dans la détection de stimuli provenant de l’environnement (extéroréception) mais peuvent également renseigner sur la position du corps et du maintien de la posture (proprioception).
V) 1. Sensilles simples
Les sensilles simples, constituées d’un poil, sont distribuées sur tout le corps de l’insecte mais sont concentrées sur des zones spécifiques en fonction de leur spécialisation.

En effet, certains d’entres eux, sensibles à tout contact avec un objet ou un substrat, sont de longueur et de sensibilité fortement variable selon les parties du corps : chez les insectes sociaux par exemple, les antennes sont riches en sensilles car ce sont des organes importants dans la communication et le toucher (voir photo 1).
D’autres poils sont spécialisés dans la détection des mouvements de l’air : ils sont situés sur la tête, le thorax et les ailes des insectes volants. Ils les renseignent sur leur vitesse, leur direction et leur position dans l’espace. Ces poils jouent donc un rôle important dans le battement des ailes et le contrôle de l’aérodynamique durant le vol (lire cet article sur le vol des insectes) (voir photo 2).

Les Orthoptères (criquets, grillons et sauterelles), et les blattes disposent quant à eux sur leurs cerques (excroissances à l’extrémité de leur abdomen) de poils longs et fins extrêmement sensibles aux déplacements et à la compression de l’air (anémorécepteurs). Ces organes peuvent ainsi détecter l’arrivée de prédateurs mais aussi les sons émis par leurs partenaires sexuels. Un infime courant d’air de 4cm/s et des sons de 30 à 1000Hz suffisent pour pour déclencher une stimulation. Les informations provenant des cerques sont envoyées puis traitées dans le ganglion abdominal terminal de l’abdomen pour former une carte tridimensionnelle de l’environnement de l’insecte.
Sur ces cerques se trouvent également des poils spécialisés, en forme de massue, dont le rôle est de détecter la gravité et de rétablir la posture de l’insecte. La pression exercée par la masse du poil sur le mécanorécepteur renseigne l’insecte sur sa posture.
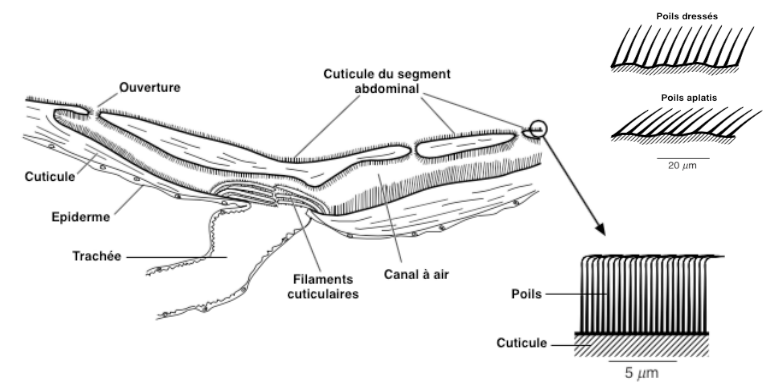
Chez la plupart des espèces aquatiques, la respiration demeure dépendante de l’oxygène de l’air. Les insectes doivent donc emprisonner une bulle d’air à la manière d’un scaphandre. Les échanges gazeux étant plus efficaces à de faibles profondeurs, il leur est important de connaître la profondeur à laquelle ils évoluent. Une punaise du genre Apheilocheirus a développé une adaptation particulière : elle présente au niveau de son deuxième segment abdominal, une invagination cuticulaire où se trouvent de nombreux petits poils hydrofuges, inclinés à 30° par rapport à la cuticule. Avec la profondeur, la pression exercée sur ces poils augmentent et l’intensité de la flexion à leur base renseigne la punaise sur sa profondeur (voir figure 4).
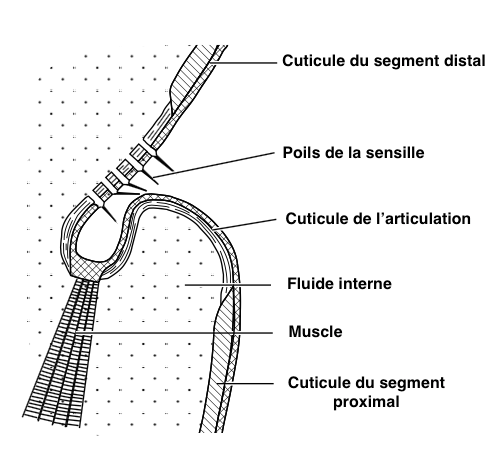
Enfin, certains poils sont disposés en rangées ou en petits amas au niveau des articulations des pattes, des antennes, des ailes et de la tête par exemple. Leur rôle est de déterminer et de renseigner l’insecte sur la position spatiale de chacune des parties de son corps afin de contrôler et de coordonner l’ensemble de ses mouvements (comme les mouvements des pattes durant la marche). Ces poils spécialisés sont appelés des sensilles trichoïdes.
La configuration spatiale particulière de ces sensilles : nombre, longueur et espacement, fournit des informations sur l’angle que forment les deux éléments de l’articulation et la vitesse à laquelle ils se déplacent entre eux (voir figure 5).
Par exemple, chez les sauterelles, on le sait, les pattes postérieures servent au saut. Pour que celui-ci soit déclenché, le fémur et le tibia doivent être repliés l’un contre l’autre et cette information est livrée à l’insecte par la stimulation de poils spécifiques. D’autres sensilles interviennent dans la régulation de la fréquence de frottements des organes stridulations à l’origine des chants sexuels.
Ches les mouches, la position de la tête par rapport au thorax durant le vol est contrôlée par des sensilles trichoïdes au niveau du cou, qui permettent à l’insecte de maintenir sa tête horizontale lors des manoeuvres.
Des sensilles trichoïdes ont pour objectif d’informer aux fourmis l’inclinaison de la pente du terrain sur lequel elles se déplacent.
V) 2. Sensilles campaniformes
Ces mécanorécepteurs sont sensibles à toute compression ou décompression se produisant à la surface de la cuticule. Leur configuration et leur orientation permettent à l’insecte d’être sensible à des forces spécifiques et d’en connaître la direction.
Ainsi, les sensilles campaniformes sont orientées perpendiculairement à la ligne de force sur les pattes : pression s’exerçant sur la cuticule par le poids de l’insecte. Elles sont notamment stimulées lorsque la patte touche le sol. L’intensité de compression renseigne l’insecte sur une éventuelle surcharge qui entraine aussitôt un mécanisme réflexe compensatoire au niveau de la hanche afin de maintenir une posture horizontale. Un réseau nerveux, reliant les sensilles campaniformes aux muscles, crée ainsi un système de contrôle à rétroaction positive ou négative selon les cas (voir figure 6).
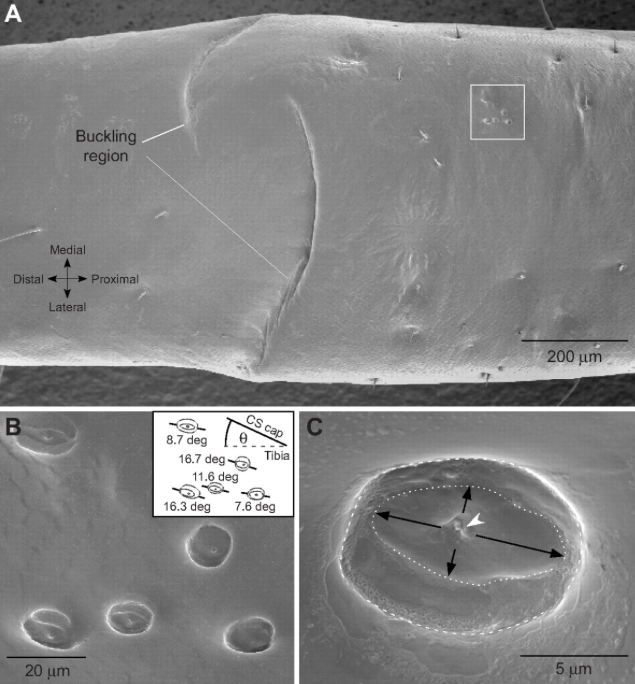
Sur les ailes, ces sensilles s’observent au niveau des nervures et des haltères (chez les mouches) et jouent un rôle important dans le contrôle du vol et la stabilité de l’insecte en détectant toute modification de la forme de l’aile, sur sa position et sa vitesse (lire les articles sur le vol des insectes : Anatomie et Aérodynamique).
Les informations provenant des sensilles campaniformes sont directement transmises aux motoneurones du thorax qui modifient la cinétique (la vitesse), la cinématique (le mouvement) du battement des ailes et donc influent sur la vitesse et la direction du vol de l’insecte.
Les insectes sociaux et les espèces devant faire appel à un sens de la communication et du toucher développé possèdent de fortes populations de sensilles campaniformes sur leurs antennes.
Une importante partie de ces signaux sont acheminés au système nerveux central (cerveau) par un réseau complexe et hautement ordonné d’axones sensoriels. Les mécanismes de traitement de l’information et la genèse des réponses adaptées (comportementales et physiologiques) feront l’objet de prochains articles.
Source :
– The Insects : Structure & Function (5ème édition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
– Keil T.A. (1997) : Functional morphology of insect mechanoreceptors – Microscopy Research and Technique, 39:506-531 (lien)
– Encyclopedia of Insects : Vincent H. RESH & Ring T. CARDE, Academic Press-2009
– Interactions Insectes-Plantes : N. Sauvion, P.A. Calatayud, D. Thiery et F. Marion-Poll – Editions Quae : 2013
Recommandation d’ouvrages sur cette thématique
– Evolution of the Insects (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
– The Insects: An Outline of Entomology (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
– The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
– Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
– Interactions insectes-plantes (N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae : 2013)
<img src= »http://wms-eu.amazon-adsystem.com/20070822/FR/img/noscript.gif?tag=passioentomo-21&linkId=3LHYKSRXQKZ3KY5I » alt= » » /><br />