Sensory receptors in insects: mechanoreceptors – Part 1: cuticular mechanoreceptors
First part : cuticular mechanoreceptors
Alike all arthropods, insects have an external skeleton, the exoskeleton, consisting of chitin and forming the cuticle (lire th article). Its insulating properties imply that the cuticle creates a barrier to the reception of stimuli from the external environment to the internal physiological environment. In order to circumvent this problem, evolution has given rise to a whole range of structures designed to transmit a wide range of types of stimuli to the insect, enabling it to grasp its environment effectively and accurately and to respond favourably to any modification by an adapted behaviour.
Among these sensory structures, some are sensitive to mechanical distortions (mechanoreceptors object of this article), others to light (photoreceptors) and others to chemical components (checeptors or chemocheceptors).
I) General
Mechanoreceptors are classified into three main categories (Table I)
| Categories | Anatomical position | Types | Functions |
| Extérorécepteurs Récepteurs cuticulaires | Cuticle surface | Senseilles simples | Touch, pressure variation (air or water), gravity detection, proprioception* |
| Campaniforms | Cup pressure variation | ||
| Interreceptors Intradermal or subcuticular receptors (click here) | Derm | Chordotonal organs | Pressure variation (air, water), gravity detection, proprioception* |
| Property receivers | Internal fabrics | No precise structure | Proprioception* |
| * Proprioception : perception of the position of the different parts of the body | |||
Featuring amazing and multiple properties, they can intervene in the sense of touch when stimulated by contact with an object or substrate, in the sense of hearing by stimulation of air or water and also in the control of posture and movement when excited by movements of parts of the insect’s body.
Two types of cuticular mechanoreceptors occur in insects: single sensils (hair on the surface of the cuticle) and campaniform sensils, dome-shaped structure where the cuticle is thin.
Despite a different structure, the cellular organization of the two types of senses is similar ( see corresponding paragraph).
II) Structural description
II) 1. Simple senses
Most insect body hairs are isolated or grouped mechanoreceptors. Depending on the species and their function, these senses, of variable length, may or may not end in a peak (and are then called trichobotries). The end of other senses end in the shape of a small club: they intervene in the detection of the magnetic field.
Evolution has allowed the appearance of as many variants as there are functions.
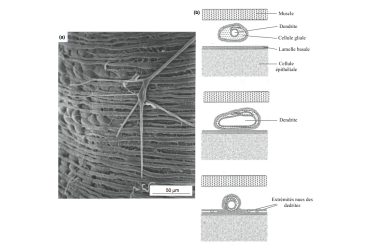
caption id= »attachment_1798″ align= »alignright » width= »350″] Figure 1 : Cross section of a simple direction (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p739 – Modified by B. GILLES)[/caption]
Figure 1 : Cross section of a simple direction (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p739 – Modified by B. GILLES)[/caption]
The anatomical structure of the simple senses is identical whatever the type (see figure 1). The sense is an outgrowth of the exocuticle whose base is composed of thin chitin thus guaranteeing a certain flexibility and movement of the hair. Elastic suspension fibres placed at this base allow a return to the initial position after a displacement. The sense rests on a partition that separates it from a cavity: the cavity of the lymphatic receptor.
This configuration is sometimes specialized to allow the direction to detect a movement in a given direction directional sensitivity.
II) 2. Campaniform senses
These oval structures, with a diameter between 5 and 30µm, consist of two cuticular layers: a thin outer layer and a fibrous and flexible inner layer. Sometimes, an additional spongy layer is inserted between these layers ( see figure 2).
caption id= »attachment_1803″ align= »alignleft » width= »350″] Figure 2 : Cross-section of a campaniform direction (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p740 – Modified by B. GILLES)[/caption]
Figure 2 : Cross-section of a campaniform direction (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p740 – Modified by B. GILLES)[/caption]
Campaniform senses are distributed over the entire surface of the insect’s body, especially in joints and areas subject to significant mechanical stress such as shins, mouth parts and dumbbells of diptera (lire cet article et celui-ci).
Many variants exist within the same individual depending on the function. Thus, in flies of the genus Calliphora, there are nearly 1200 bell-shaped senses divided into nine types: 36 per leg, 140 at the base of each wing and 340 on each dumbbell.
III) Cell description
The simple and bell-shaped senses are linked to a complex of several epidermal cells composed of a sensory nerve cell (neuron) and two or three support cells.
- Le sensory neuron, locatable at its particularly large nucleus, is attached to the base of the hair of the simple direction and inserted into the two cuticular layers of the bell-shaped direction by a single unbranched dendrite and surrounded by a sheath ( see figure 1 & 2).
- The support cells are most often three: thecogene (closest to the neuron) secretes the sheath of the dendrite, trichogen secretes the cuticle of the hair and tormogen (outermost) secretes the septum of the lymphatic cavity of the receptor ( see Figure 1). In some species, the latter can disappear once its role has expired.
Dendrite consists of three parts: a distal part containing numerous mitochondria and other cellular organelles, a ciliated narrowing zone at the cuticle (endocuticle) and, in its proximal part, a tubular body comprising a large number of microtubules ( tubulin polymer) which make it extremely sensitive to variations in shape ( see Figure 2).
IV) Functioning
The operation of cuticular mechanoreceptors involves three successive processes: 1) coupling receptor/generator potential and 3) action potential.
IV) 1. The coupling
The coupling is the mechanism that collects the mechanical energy captured by the hair or the campaniform direction.
When the hair of the sense comes into contact with an object or substrate, a bending force is exerted at its base, such as a lever : it generates an amplification of movement and causes a mechanical distortion at the level of the tubular body of the sensory neuron.
The same principle is observed in the campaniform direction where the coupling is carried out by the compression or decompression of the cuticle at this place. Some tubular bodies have a fan shape which gives directional sensitivity to the bell-shaped direction.
Multiple types of senses are present on the insect’s body: up to 12 types have been recorded on one cockroach leg. Their variations in shape, size and orientation provide the insect with a great diversity of information.
VI) 2. Transduction & potential generator
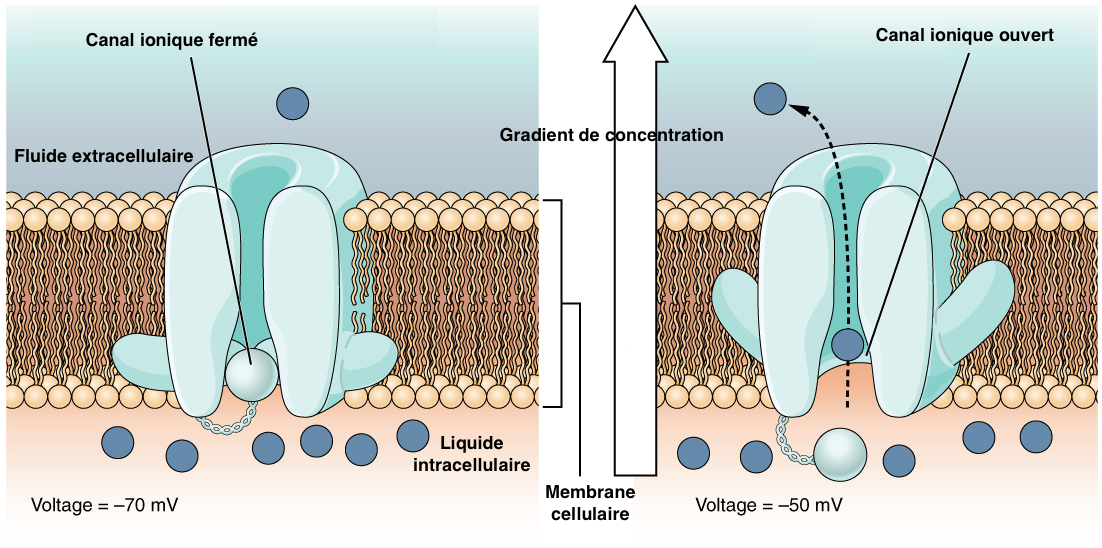
This process corresponds to the translation of mechanical energy into electrical energy at the level of the tubular body of the sensory nerve cell. The electrical energy thus translated is the potential generator (PG), also called receiver potential.
Mechanical action on the hair or the campaniform direction causes a deformation of the tubular body, leading to the opening of ionic channels (most often potassium channels) on the surface of the plasma membrane. This ion exchange creates a depolarization (transient from a negative value to a positive value on the surface of a cell) and thus an electric current ( see figure 3).
Sensitivity is such that a tubular body distortion of barely 5nm causes a PG!
IV) 3. Potential for action
The potential for action (PA) follows depolarization of the plasma membrane above a certain threshold value. Also called nervous swelling, it corresponds to the transmission of this depolarization along the dendrite and neuron, and thus the sending of the nervous message to other neurons through the body.
The process consists in a successive opening/closing of the ion channels modifying the electrical charge of the membrane (see videos below). Thus, the membrane potential increases from a value of -70mV to +30mV then returns to its initial value to be activated again. This cycle takes an average of 2 milliseconds.
Each type of mechanoreceptor has a specific depolarization and AP threshold. The generation of a PA depends on the force acting on the mechanoreceptor, the coupling sensitivity, the nature of the ion channels, the intensity and frequency of depolarization.
The sensitivity and reaction following a mechanoreceptor stimulation are thus very variable.
In addition, the response to a stimulus can be of two types: phasic or tonic.
An adequate mechanical pressure on the mechanoreceptor leads to the appearance of an action potential (AP). A response is said to be phasic if maintaining this pressure does not cause a new PA. This type of answer concerns in particular the hair of the simple senses.
Conversely, if several APs are emitted during the entire mechanical pressure phase, the response will be tonic and typical of the campaniform senses response. Some campaniform senses can trigger either tonic or phasic responses depending on the type, frequency and intensity of the stimulus.
The treatment and activation of action potentials are regulated by numerous biochemical and electrical processes. The insect’s response to a range of stimuli results from the analysis of all stimuli, their intensity, frequency and amplitude at all times.
V) Examples of the role played by cuticular mechanoreceptors
Simple hair plays a role in detecting environmental stimuli (extéroréception) but can also provide information on body position and posture maintenance (proprioception).
V) 1. Senseilles simples
The simple senses, consisting of a hair, are distributed over the entire body of the insect but are concentrated on specific areas according to their specialization.
caption id= »attachment_1824″ align= »alignright » width= »380″] Photo 1 : Senses of touch on a cockroach antenna in Madagascar (Source : Photo by Jaspen Nance – Flickr.com)[/caption] caption id= »attachment_1807″ align= »aligncenter » width= »517″] Figure 3 : Opening of the ion channel and ion exchange at the surface of the plasma membrane of the nerve cell (Source : Wikipedia – Modified by B. GILLES)[/caption]
Photo 1 : Senses of touch on a cockroach antenna in Madagascar (Source : Photo by Jaspen Nance – Flickr.com)[/caption] caption id= »attachment_1807″ align= »aligncenter » width= »517″] Figure 3 : Opening of the ion channel and ion exchange at the surface of the plasma membrane of the nerve cell (Source : Wikipedia – Modified by B. GILLES)[/caption]
IV) 3. Potential for action
The potential for action (PA) follows depolarization of the plasma membrane above a certain threshold value. Also called nervous swelling, it corresponds to the transmission of this depolarization along the dendrite and neuron, and thus the sending of the nervous message to other neurons through the body.
The process consists in a successive opening/closing of the ion channels modifying the electrical charge of the membrane (see videos below). Thus, the membrane potential increases from a value of -70mV to +30mV then returns to its initial value to be activated again. This cycle takes an average of 2 milliseconds.
Each type of mechanoreceptor has a specific depolarization and AP threshold. The generation of a PA depends on the force acting on the mechanoreceptor, the coupling sensitivity, the nature of the ion channels, the intensity and frequency of depolarization.
The sensitivity and reaction following a mechanoreceptor stimulation are thus very variable.
In addition, the response to a stimulus can be of two types: phasic or tonic.
An adequate mechanical pressure on the mechanoreceptor leads to the appearance of an action potential (AP). A response is said to be phasic if maintaining this pressure does not cause a new PA. This type of answer concerns in particular the hair of the simple senses.
Conversely, if several APs are emitted during the entire mechanical pressure phase, the response will be tonic and typical of the campaniform senses response. Some campaniform senses can trigger either tonic or phasic responses depending on the type, frequency and intensity of the stimulus.
The treatment and activation of action potentials are regulated by numerous biochemical and electrical processes. The insect’s response to a range of stimuli results from the analysis of all stimuli, their intensity, frequency and amplitude at all times.
V) Examples of the role played by cuticular mechanoreceptors
Simple hair plays a role in detecting environmental stimuli (extéroréception) but can also provide information on body position and posture maintenance (proprioception).
V) 1. Senseilles simples
The simple senses, consisting of a hair, are distributed over the entire body of the insect but are concentrated on specific areas according to their specialization.
caption id= »attachment_1824″ align= »alignright » width= »380″] Photo 1 : Senses of touch on a cockroach antenna in Madagascar (Source : Photo by Jaspen Nance – Flickr.com)[/caption]
In fact, some of them, sensitive to any contact with an object or a substrate, are of length and sensitivity strongly variable according to the parts of the body: in the social insects for example, the antennas are rich in directions because they are important organs in the communication and the touch ( see photo 1).
Other hairs are specialized in detecting air movements: they are located on the head, thorax and wings of flying insects. They inform them about their speed, direction and position in space. These hairs therefore play an important role in wing flapping and aerodynamic control during flight (lire this article on insect flight) (see photo 2).
caption id= »attachment_1825″ align= »aligncenter » width= »594″] Photo 2 : Example of senses and their role on a fly like Tephritis (Tephritidae) (Source : ©Photo B. GILLES)[/caption]
Photo 2 : Example of senses and their role on a fly like Tephritis (Tephritidae) (Source : ©Photo B. GILLES)[/caption]
Orthopterans (locusts, crickets and grasshoppers), and cockroaches have long, fine hairs on their cerci (extrusions at the tip of their abdomen) that are extremely sensitive to air movement and compression (anemoreceptors). These organs can thus detect the arrival of predators but also the sounds emitted by their sexual partners. A very small air current of 4cm/s and sounds from 30 to 1000Hz are enough to trigger a stimulation. The information from the cherries is sent and processed into the terminal abdominal node of the abdomen to form a three-dimensional map of the insect environment.
On these cerques are also specialized hairs, in the shape of a club, whose role is to detect the gravity and restore the posture of the insect. The pressure exerted by the mass of hair on the mechanoreceptor informs the insect on its posture.
In most aquatic species, breathing remains dependent on oxygen in the air. Insects must therefore trap an air bubble like a suit. As gas exchanges are more efficient at shallow depths, it is important for them to know the depth at which they evolve. A bug of the genus Apheilocheirus has developed a particular adaptation: it presents at the level of its second abdominal segment, a cuticular invagination where numerous small water-repellent hairs are found, inclined at 30° with respect to the cuticle. With depth, the pressure exerted on these hairs increases and the intensity of flexion at their base informs the chinch bug about its depth ( see figure 4).
caption id= »attachment_1833″ align= »aligncenter » width= »632″] Figure 4 : Pressure detection device for chinch bugs of the genus Aphelocheirus (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p536 – Modified by B. GILLES)[/caption]
Figure 4 : Pressure detection device for chinch bugs of the genus Aphelocheirus (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p536 – Modified by B. GILLES)[/caption]
Finally, some hairs are arranged in rows or in small clusters at the joints of the legs, antennas, wings and head for example. Their role is to determine and inform the insect about the spatial position of each part of its body in order to control and coordinate all of its movements (such as leg movements during walking). These specialized hairs are called sensils trichoid.
The particular spatial configuration of these meanings: number, length and spacing, provides information on the angle formed by the two elements of the joint and the speed at which they move between them ( see figure 5).
caption id= »attachment_1834″ align= »alignright » width= »310″] Figure 5 : Trichoid direction diagram at a joint (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p746 – Modified by B. GILLES)[/caption]
Figure 5 : Trichoid direction diagram at a joint (Source : The Insects : structure & function – R.F. Chapman – 5th edition, 2013 – p746 – Modified by B. GILLES)[/caption]
For example, in grasshoppers, we know, the hind legs are used for jumping. For this to be triggered, the femur and tibia must be folded together and this information is delivered to the insect by stimulating specific hairs. Other senses are involved in the regulation of the frequency of friction of the chirping organs at the origin of sexual songs.
Flies, the position of the head relative to the thorax during flight is controlled by trichoid senses at the neck, which allow the insect to maintain its head horizontal during manoeuvres.
Trichoid senses aim to inform ants of the slope of the terrain they are moving on.
V) 2. Campaniform senses
These mechanoreceptors are sensitive to any compression or decompression occurring on the cuticle surface. Their configuration and orientation allow the insect to be sensitive to specific forces and to know their direction.
In this way, the campaniform senses are oriented perpendicularly to the line of force on the legs : pressure exerted on the cuticle by the weight of the insect. They are particularly stimulated when the leg touches the ground. The intensity of compression informs the insect about a possible overload which immediately leads to a compensatory reflex mechanism at the hip level in order to maintain a horizontal posture. A nervous network, connecting the campaniform senses to the muscles, thus creates a control system with positive or negative feedback depending on the case ( see figure 6).
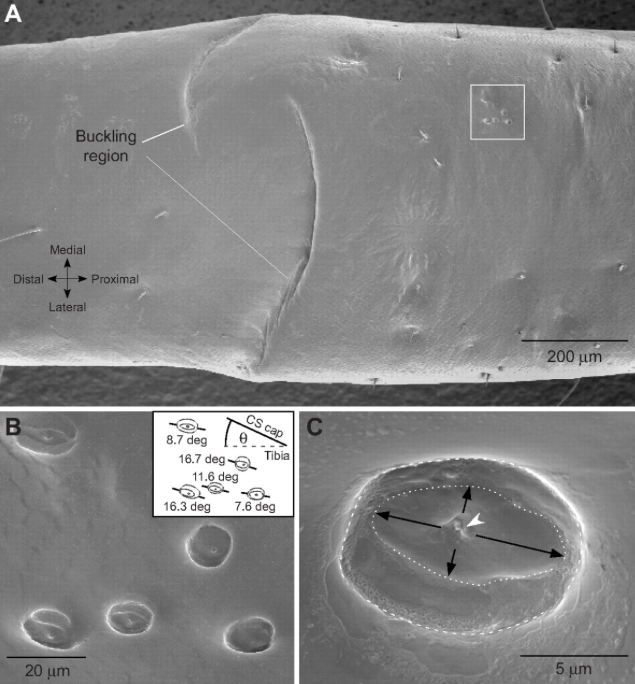
On the wings, these senses are observed on the ribs and dumbbells (in flies) and play an important role in the control of the flight and the stability of the insect by detecting any modification of the shape of the wing, on its position and its speed (read the articles on the flight of insects Anatomie et Aérodynamique).
Information from the campaniform senses is directly transmitted to the thorax motor neurons which modify the kinetics (speed), kinematics (movement) of wing flapping and thus influence the speed and direction of insect flight.
Social insects and species requiring a developed sense of communication and touch have strong populations of bell-shaped senses on their antennas.
A significant portion of these signals are sent to the central nervous system (brain) through a complex and highly ordered network of sensory axons. Information processing mechanisms and the genesis of adapted responses (behavioural and physiological) will be the subject of future articles.
Source :
– The Insects : Structure & Function (5th edition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
– Keil T.A. (1997) : Functional morphology of insect mechanoreceptors – Microscopy Research and Technique, 39:506-531 (lien)
– Encyclopedia of Insects : Vincent H. RESH & Ring T. CARDE, Academic Press-2009
– Insect-Plant Interactions : N. Sauvion, P.A. Calatayud, D. Thiery and F. Marion-Poll – Editions Quae : 2013
Recommendation of books on this theme
>
– Evolution of the Insects (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – May 16, 2005)
– The Insects: An Outline of Entomology (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5th edition : 6 August 2014)
– The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5th edition : 12 November 2012)
– Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2nd edition : 22 July 2009)
– Interactions insectes-plantes (N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae : 2013)