Parthenogenesis in insects: synthesis
by Benoit GILLES
In order for them to multiply, nature has endowed living organisms with a wide diversity of reproductive systems. In insects, one of these strategies is parthenogenesis. This is based on the development of individuals from unfertilized gametes, thus without the need for fertilization. This strategy is interesting for more than one reason: it is evidenced by the declination of a multitude of forms (thelytokia, arrhenotokia and deuterotokia) and its appearance on multiple occasions during evolution, within unrelated and phylogenetically distant taxa and species.
Description of the different types of parthenogenesis
I) Thelytokia
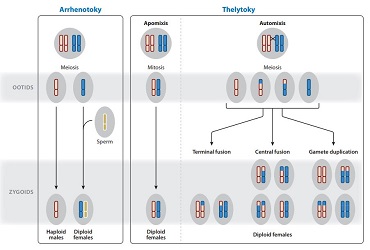
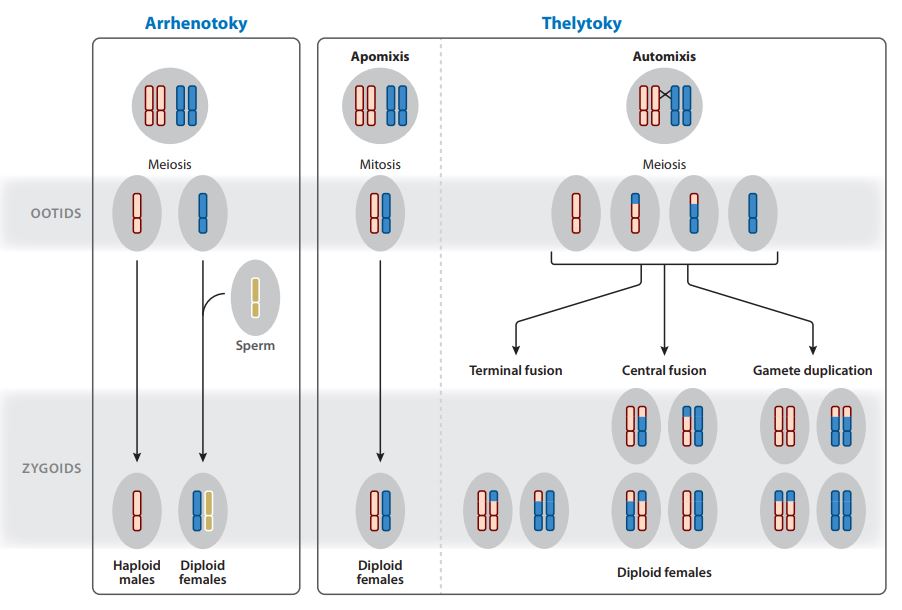
Corresponding to parthenogenesis sensu stricto, thelytokia is characterized by the fact that all the unfertilized eggs emitted by a female insect in turn only produce a female diploid progeny.
Thelytoic parthenogenesis can result in two distinct genetic systems: one based on mitosis (apomixis) and the other on meiosis (automixis).
- L’apomixie is the simplest system: female offspring are genetically similar to the mother, the absence of meiosis prevents any chromosomal rearrangement (genetic mixing). Mother and daughter can thus be considered as clones
- L’automixie involves the meiosis process, diploidy is restored without the contribution of a gamete (without fertilization). This process can be carried out in several ways: 1) duplication of the genome before meiosis, creating a cell with 4n chromosomes (4 sets of chromosomes); 2) fusion of the two nuclei during meiosis; 3) a haploid nucleus resulting from meiosis is duplicated by mitosis to fuse again with itself
II) Arrhenotokism
In arrhenotokic parthenogenesis, progeny from unfertilized eggs consist only of male individuals (opposite system to thelytokia). There are two distinct genetic systems: 1) haplodiploid, males are haploid (1 set of chromosomes) and females are diploid (2 sets of chromosomes); 2) diploid, males and females are derived from diploid eggs as for thelycan parthenesis.
III) Deuterotokia
Deutherotokic parthenogenesis produces unfertilized eggs from both males and females.
Appearance of parthenogenesis
A parthenogenetic system can appear in various ways. The most singular case is based on the interaction between certain groups of insects and endosymbiotic bacteria such as Wolbachia, Rickettsia and Cardinium – the most widely represented being Wolbachia pipientis – causing cytoplasmic incompatibilities causing either the cessation of the development of diploid embryos, or the thelytokia, or the feminization of males, their death. These bacteria often play a role in sex conversion and modify the sex ratio in favour of females.
Many insect species are monosexed (their population is composed of only one sex). These species occur in isolated environments such as islands or high-altitude regions. They perform a geographic parthenogenesis, demonstrating the adaptive power of this type of reproduction.
>Parthenogenesis in hemimetaboles (or heterometaboles)
Hemimetazole insects federate species whose metamorphosis is said to be incomplete: the adult stage is reached gradually during the development cycle by successive moulting, the larvae possessing most of the attributes of adults, except sexual and parasexual organs (link article).
Odonata (dragonflies) : only the species Ischnura hastata (Caenagrionidae), found in the Azores, has been described as parthenogenetic (thelytoic type).
Orthoptera (locusts, crickets and grasshoppers) : the species Locusta migratoria and Schistocerca gregaria have the ability to spontaneously generate female offspring from unfertilized eggs: thycoparthenogenesis. Haploid gametes become diploid gradually during development. For Loxoblemmus frontalis, the only species in the Gryllidae family to practice parthenogenesis, thelytokia is induced by the presence of bacteria Wolbachia. It is without intervention of Wolbachia in grasshoppers Saga pedo (Tettigonidae).

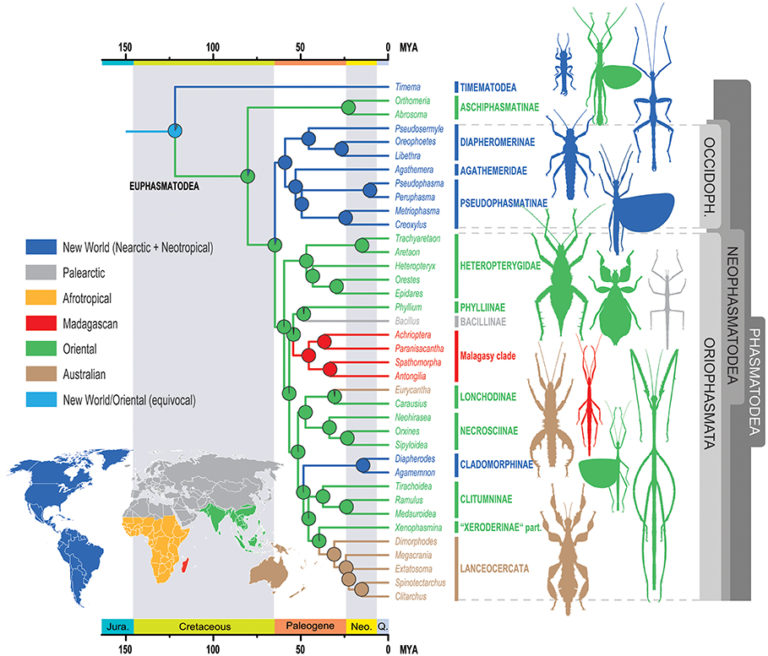
Phasmatodea (Phasms) : parthenogenesis is quite common. For example, the genus Timema, endemic to California, is composed of 5 species all related and descended from the same lineage. However, rare fertile males could be collected, their presence being estimated at less than 0.2% of the population. The sexual determination of phasms is based on a system where individuals carrying a pair of autosome chromosomes (XX) are female and those with only one copy of the X are male (XO), (in humans the system is XX-XY), males develop via spontaneous loss of the X chromosome during oogenesis. Other species also reproduce only by parthenogenesis such as Bacillus rossius and Clonopsis gallica (species found in southern France).
Blattoptera (formerly Isoptera) (termites) : several groups optionally use thelycan parthenogenesis through a ploidy restoration process similar to that found in Reticulitermes speratus and R. virginicus (automiscie). This type of termites is known to harbour Wolbachia, whether the colonies are parthenogenetic or bisexual, suggesting that there is no correlation between the two reproductive modes.
Mantodea (mantes) (lien) : only two species, Miomantis savignyi and Bruneria borealis, carry out parthenogenesis (thelytoque), mandatory parthenogenesis in B. borealis.
Hemiptera (cicadas, aphids and bedbugs) : this order offers an abundant diversity of parthenogenetic systems within a large number of families such as Aclerdidae, Diaspididae, Aphididae or Anthocoridae. For example, several species of Fulgores (Delphacidae) belonging to the genera Delphacodes and Ribautodelphax use thelytokia and pseudogamy (females mate with males but the offspring are entirely female). The intervention of Wolbachia in the parthenogenesis process has been demonstrated in Delphacodes kuscheli but not in other species. A multitude of forms of parthenogenesis occur between species of the Coccidae and Diapsididae families: arrhenotokism with males that may be diploid or haploid, and deuterotokism.
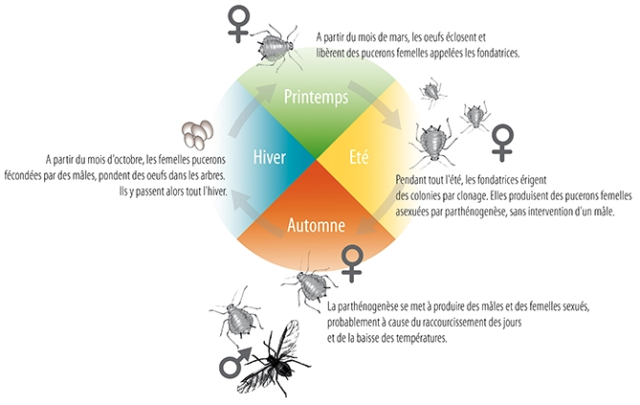
Some aphid species (Aphidimorpha) have a cyclic reproductive mode, changing from bisexual to parthenogenetic depending on the season. Thus, in spring, a female aphid (founder) multiplies by parthenogenesis in order to rapidly colonize the environment, then, in autumn, reproduction becomes sexual, resulting in the production of fertilized eggs that overwinter in the vegetation.
With regard to « real » bedbugs, there are no cases of parthenogenesis revealed. However, two species have optional use of telytokism: Calliodis maculipennis (neotropical species of the Anthociridae family) and Campyloneura virgula (Miridae). Surprisingly, a parthenogenetic population of C. maculipennis located on the island of Trinidad has been discovered, while the populations of the mainland, Mexico and Guyana, are gendered.
Life cycle of the aphid (english)
Parthenogenesis in holometabolas
The holometabol group characterizes species using a so-called complete metamorphosis phase where the passage to adulthood requires a chrysalis (butterfly) or pupa (flies) phase during which the larvae are totally transformed (lien article).
Hollo-metabolas make up the vast majority of insect species diversity: 800,000 species divided into 11 orders such as Diptera, Coleoptera, Lepidoptera or Hymenoptera.
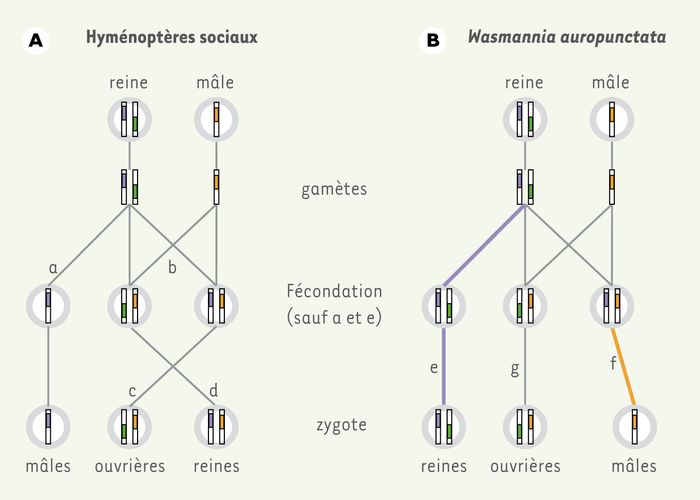
The Hymenoptera (wasps, bees, ants), with nearly 150,000 species described, constitutes one of the most diversified insect orders. Parthenogenesis is common to all species of the order, thus encompassing the greatest diversity of types of parthenogenesis. The most common and ancestral is arrhenotokism combined with haplodiploidy. Females thus have the ability to fertilize or not their eggs and can adjust the sex ratio of their offspring: a diploid egg will give a sterile female or worker and a haploid egg a male (see illustration below). However, mechanisms may vary between taxa.
The transition from arrhenotopic to thelytoic parthenogenesis is relatively frequent due to the absence of a sexual chromosome making it possible to restore diploidy through an apomictic or automictic process. For example, the species Diplolepis eglanteria (Cynipidae) is a small wasp that uses apomixis, and Apis mellifera capensis a, on the other hand, uses self-mixing.

Cataglyphis hispanica, Paratrechina longicornis, Vollenhovia emeyri and Wasmannia auropunctata are derived from sexual reproduction while the new queens come from parthenogenetic thelycan eggs. The study of the reproductive mode of Wasmannia auropunctata revealed a unique case of a dual parthenogenetic system (arrhenotokic and thelytok). It was discovered in 2005 that males, derived from fertilized eggs, expressed only the paternal genome, with the maternal genome disappearing (except the mitochondrial genome) through a mechanism that remains partly unknown. This process indicates that male offspring are cloned (see illustration opposite).
So far as these workers are sterile, this sexual reproduction does not lead to the mixing of male and female genomes in the next generation. This lack of gene flow between males and females leads to genetic differentiation and separate evolution of the two genomes. This raises the question as to the classification of a possible distinction between these two sexes as two distinct species, one of which would only consist of males! In addition, males can be considered as parasites exploiting females, where the production of sterile workers ensures the protection and supply of the colony.
At 30% of parasitoid wasp species (article link) Cynipidae and Chalcidoidea, thelytokia is caused by endosymbiotic microorganisms of the genus Wolbachia, Cardinium and Rickettsia.
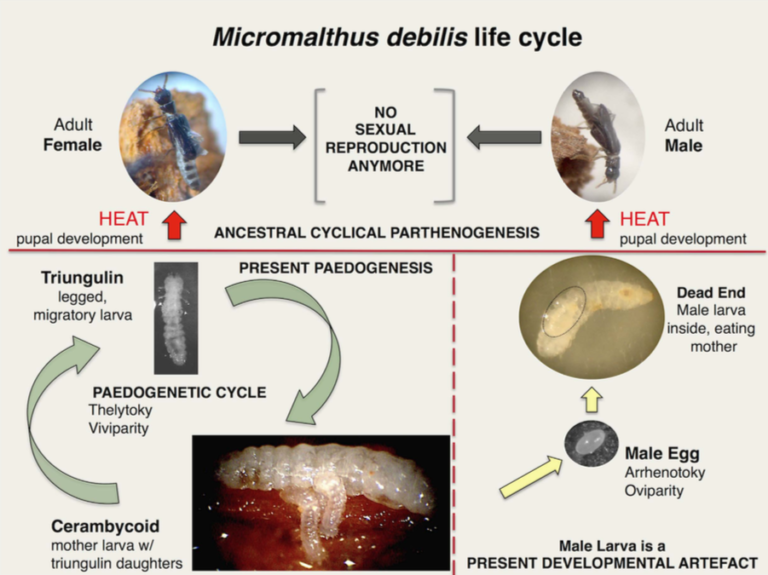
The order of Coleoptera accounts for 30% of insect species, or 380,000 species of which only 600 species (20 families) are parthenogenetic. Thelytokia is more widespread than arrhenotokia: it is found in Alexiidae, Anobiidae, Cerambycidae, Dermestidae, Elateridae, Hydrophiliidae, Passalidae, Sphindidae or Staphylinidae, among others. The case of Micromalthus debilis is unique in its kind, the thelytokia is pushed to the extreme because the sterile males have almost disappeared from the populations (lire this article).
So, the species Reesa vesopulae (Dermestidae), present throughout the nearctic region, is strictly parthenogenetic, as are the majority of North American populations of Cis fuscipes (Ciidae) and Aelus mellillus (Elateridae). In Chrysomelidae, the species Bromius obscurus is represented by diploid bisexual populations in North America and other apomictic triploids in Europe. The two European weevil species (Curculionidae), Polydrus mollis and Otiorhynchus scaber are mainly thelytoics, although some populations are bisexual diploids in small localities.
In Lepidoptera, despite more than 170,000 species described, only two dozen species practice parthenogenesis. These species are mainly Lymantriidae and Psychidae (11 species). There is a gendered form of Dahlica triquetrella in Central Europe, while several diploid and tetraploid thelytok populations are widespread throughout Europe and North America.
Flies (Dipteran Orders), parthenogenesis has appeared in at least 11 families with more than 150 000 species : Chironomidae, Hybotidae, Agromyzidae, Cecidomyiidae, Psychodidae, Sciaridae, Ctenostylidae, Lonchopteridae, Simuliidae, Ceratopogonidae and Chamaemyliidae. Half of the Chironomidae taxa studied appear parthenogenetic. This strategy seems to have been selected to survive extreme environmental conditions, particularly cold (altitude and latitude), such as Eretmoptera murphyi (Antarctica) and Micropsectra sedna (Canada). Others, such as Cladotanytarsus aeiparthenus and Paratanytarsus grimmii, live in polluted or acidic waters.
Troglocladius hajdi and Lymnophyes minimus living in the Gough and Nightingale Islands south of the Atlantic Ocean, or Monopelopia caraguata, Phtytelmatocladius delarosai and Polypedilum parthenogeneticum which live in small puddles (water accumulated in the leaf axil, trunk cavity, etc.)) located on terrestrial plants: phytotelme, from ancient Greek phyto – plant telma – pond.