Récepteurs sensoriels chez les insectes : les Mécanorécepteurs – Deuxième partie : les organes chordotonaux
Deuxième partie : les organes chordotonaux
Comme l’ensemble des arthropodes, les insectes ont un squelette externe, l’exosquelette, constitué de chitine et formant la cuticule (lire cet article). Ses propriétés isolantes impliquent que la cuticule crée une barrière à la réception des stimuli de l’environnement extérieur vers le milieu physiologique interne. Afin de contourner cette problématique, l’évolution a fait surgir tout un ensemble de structures destinées à transmettre une large gamme de types de stimuli à l’insecte qui lui permettent d’appréhender de manière efficace et précise son environnement et de répondre favorablement à toute modification par un comportement adapté.
Parmi ces structures sensorielles, certaines sont sensibles aux distorsions mécaniques (mécanorécepteurs : objet de cette article), d’autres à la lumière (photorécepteurs) et d’autres encore à des composants chimiques (chémorécepteurs ou chimiorécepteurs).
I) Généralités
Les mécanorécepteurs sont classés en trois grandes catégories (Tableau I) :
| Catégories | Position anatomique | Types | Fonctions |
| Extérorécepteurs Récepteurs cuticulaires (cliquez ici) | Surface de la cuticule | Sensilles simples | Toucher, variation de pression (air ou eau), détection de la gravité, proprioception* |
| Sensilles campaniformes | Variation de pression sur la cuticule | ||
| Intérorécepteurs Récepteurs intradermiques ou subcuticulaires | Derme | Organes chordotonaux | Variation de pression (air, eau), détection de la gravité, proprioception* |
| Propriorécepteurs | Tissus internes | Pas de structure précise | Proprioception* |
| * Proprioception : perception de la position des différentes parties du corps | |||
Dotés de propriétés étonnantes et multiples, ils peuvent intervenir dans le sens du toucher lorsqu’ils sont stimulés par le contact d’un objet ou un substrat, dans le sens de l’ouïe par une stimulation de l’air ou de l’eau et aussi dans le contrôle de la posture et des mouvements quand ils sont excités par des déplacements de parties du corps de l’insecte.
II) Description structurelle
Les organes chordotonaux, ou scolopophores, sont des récepteurs intradermiques agissant le plus souvent comme des propriocepteurs (perception de la position des différentes parties du corps) ou intervenant dans l’audition.
Ils sont constitués d’éléments simples appelés scolopidies, attachés à la cuticule par une ou plusieurs de leurs extrémités. Ils peuvent être solitaires ou regroupés en amas.
Chaque scolopidie est composée de 3 cellules (voir figure 1) :
- un neurone sensoriel
- une cellule enveloppante (cellule scolopale)
- une cellule d’attache
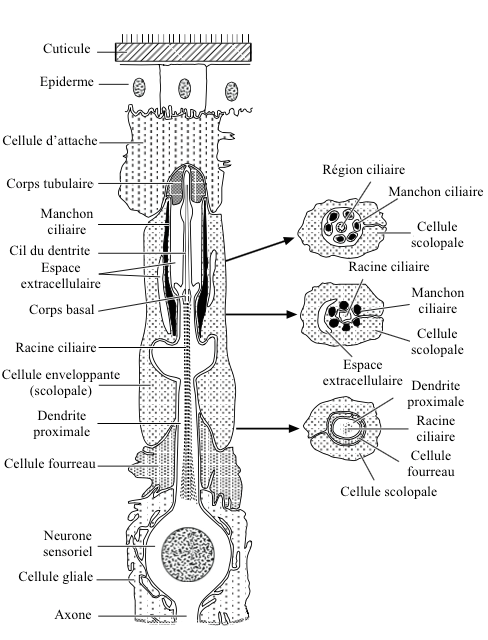
La dendrite du neurone sensoriel, recouverte par la cellule enveloppante, possède à son extrémité une extension ciliaire insérée à la cellule d’attache par un corps tubulaire.
La région ciliaire, située dans un espace extracellulaire, est rigidifiée par la présence de tiges d’active sécrétées par la cellule scolopale (structure similaire à un « manchon en forme de tonneau »). Un second espace extracellulaire, rencontré à la base du manchon, est appelé « collier ciliaire » en raison de son agencement.
A l’intérieur de la dendrite neuronale, une structure composée de 9 filaments doubles partant de la base de la région ciliaire, court jusqu’à sa partie proximale : la racine ciliaire.
Dans la plupart des cas, une seule dendrite s’insère dans le manchon ciliaire. Parfois, dans certaines scolopidies, plusieurs dendrites (jusqu’à 5) se réunissent dans le manchon ciliaire.
La cellule d’attache joue le rôle de lien entre l’organe chordotonal et l’épiderme ou d’autres types de tissus.
II) Fonctionnement
Le fonctionnement de l’organe chordotonal repose sur trois processus successifs : 1) le couplage, 2) le potentiel récepteur/générateur, 3) le potentiel d’action.
Le rapprochement de deux plaques cuticulaires (par exemple deux éléments d’une articulation) l’une de l’autre induit une modification de la structure de la scolopidie. La déformation structurelle produit un déplacement du manchon ciliaire conduisant à la flexion du cil de la dendrite. Cette action permet de recueillir l’énergie mécanique au niveau cellulaire, ce qui correspond au couplage.
La flexion du cil déclenche l’ouverture de canaux ioniques (potassiques le plus souvent) situés à la surface de la membrane cellulaire. Cet échange d’ions crée une dépolarisation (passage transitoire d’une valeur négative à une valeur positive à la surface d’une cellule) et donc d’un courant électrique. Ce mécanisme permet de converti l’énergie mécanique en énergie électrique. L’énergie ainsi produite est le potentiel générateur (PG), aussi appelé potentiel de récepteur.
Une dépolarisation au delà d’un certain seuil entraine la formation d’un potentiel d’action (PA), aussi appelé influx nerveux. Celui-ci permet la transmission de la dépolarisation le long de la dendrite et du neurone, et donc de l’envoi du message nerveux vers d’autres neurones à travers l’organisme. Le principe consiste en une ouverture/fermeture successive des canaux ioniques modifiant le potentiel électrique de la membrane.
La réponse à un stimulus peut être soit tonique, soit phasique. Elle est tonique lorsque le maintien de la flexion du cil (modification structurelle de la scolopidie) provoque l’émission de plusieurs PA successifs, au contraire, elle est dite phasique quand aucun autre PA n’est émis.
Pour en savoir davantage sur ces processus, je vous invite à lire le paragraphe correspondant dans cet article.
III) Distribution et fonction chez les insectes
Les organes chordotonaux sont présents sur tout le corps des insectes. Leur fonction est multiple : proprioception – détection de vibration du substrat, de l’air et de l’eau – détection de la gravité – audition.
Par exemple, certains de ces organes se trouvant dans la chambre génitale ont pour rôle de détecter la pression exercée par l’arrivée des ovocytes. Leur stimulation déclenche des mécanismes musculaires spécifiques permettant le reflux des spermatozoïdes de la spermatique pour réaliser la fécondation.
D’autres, situés dans le thorax, sont de grande taille (20 scolopidies) et interviennent dans le contrôle du mouvement de la tête par rapport au thorax.
A la base des ailes, ces organes sont stimulés par les forces exercées durant le vol (lire cet article).
Dans une patte d’insecte, 4 types d’organes chordotonaux, voir davantage, peuvent être présents :
- un fixé au fémur (chez les Orthoptères) dans sa partie distale, constitué d’environ 300 scolopidies
- un fixé au tibia de manière proximale : c’est l’organe subgénual
- un dans le tissu conjonctif du tibia, inséré dans l’articulation tibia-tarsienne
- un de petite taille (3 scolopidies) dans l’articulation du tarse aux prétarses
Des organes chordotonaux particuliers se situent dans les pédoncules des antennes de tous les insectes : les organes de Johnston.
IV) Types d’organes chordotonaux
IV) 1. Organes fémoraux
Il semble que tous les insectes possèdent des organes chordotonaux au niveau de l’articulation fémoraux-tibiale (FeCO). Ces organes ont particulièrement bien été étudiés.
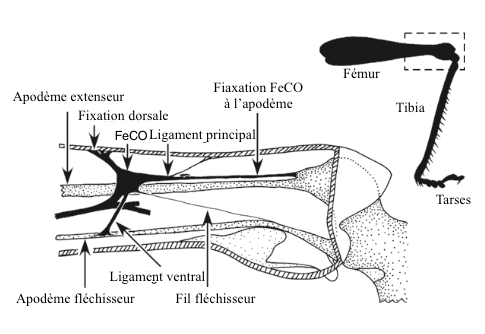
Chez les Orthoptères (criquets, sauterelles et grillons), l’organe est ancré au niveau de l’hypoderme (couche profonde du derme) du fémur où il est attaché par un ligament, et dans l’apodème (squelette externe situé à l’intérieur d’une articulation chez les arthropodes) de la tête du tibia près du muscle extenseur (voir figure 2). D’autres éléments interviennent pour maintenir la structure tridimensionnelle de l’organe. Des arrangements plus simples se rencontrent chez d’autres insectes comme les Diptères (mouches), les Lépidoptères (papillons) et les Hémiptères (punaises) où l’organe chordotonal est directement inséré dans les fibres musculaires.
Cet organe a pour rôle de renseigner l’insecte sur l’accélération, la vitesse et la position du tibia par rapport au fémur lors d’un saut ou de la marche. D’autres organes interviennent également dans le contrôle des mouvements : sensilles simples et sensilles campaniformes (lire cet article).
Les organes chordotonaux sont reliés à des arcs neuronaux réflexes situés dans les muscles extenseurs et de flexion des pattes : ils contribuent au maintien de la posture durant la marche (le vol ou la nage) et aux mouvements spécifiques accomplis par l’insecte : mouvement mimétique d’une feuille dans le vent chez le phasme par exemple.
IV) 2. Organes subgénuaux
Ces organes, situés dans les parties proximales des tibias, sont en général composés de 10 à 40 scolopidies, parfois beaucoup plus, jusqu’à 400 chez certaines guêpes parasitoïdes (qui pondent et se développent dans des oeufs et le corps d’autres insectes).
Contrairement aux organes fémoraux, ils ne sont pas associés à une articulation. Les scolopidies, regroupées en amas, sont en contact avec une trachée (espace où circule l’air pour la respiration) à une extrémité et fixées de l’autre à la cuticule par l’intermédiaire des cellules d’attache (voir figure 3).
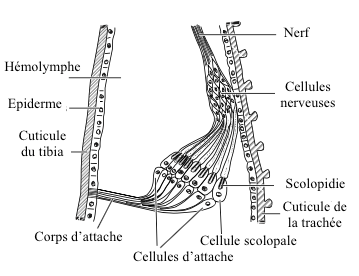
L’organe subgénual est constitué de deux parties (proximale, distale) chez de nombreux insectes comme les Odonates (libellules), les Blattodea (blattes) et les Orthoptères ; d’une seule partie distale chez les Homoptères (cigales), les Hétéroptères (punaises), les Neuroptères (chrysopes) ou encore les Coléoptères et les Diptères.
Sensibles aux vibrations du substrat, ils jouent ainsi un rôle soit dans la détection de l’arrivée d’un prédateur, soit dans la communication intra-spécifique (recherche de partenaires sexuels ou agrégation).
Ces organes sont d’une très grande sensibilité. Par exemple, chez la blatte du genre Periplaneta, un déplacement de seulement 0,2 nanomètre (0,2 milliardième de mètre!) et d’une fréquence d’1,5kHz suffit à déclencher une réponse nerveuse (PA).
Baignant dans l’hémolymphe (liquide extracellulaire chez les arthropodes), les mouvements de l’organe subgénual subissent une certaine inertie (il peut être considéré comme un piston dans un tube rempli de liquide).
Chez les guêpes parasitoïdes (Ichneuminidae), les organes chordotonaux possèdent une autre fonction : celle de rechercher les proies (hôtes) se trouvant sous les écorces des arbres.
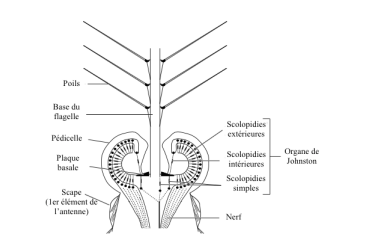
IV) 3. Organes de Johnston

Ces organes, situés dans le pédoncule antenaise de tous les insectes adultes ou sous forme plus simplifiée chez les larves, présentent la caractéristique d’être constitués d’un flagelle qui est incorporé à un pédicelle dont la base comprend de très nombreux scolopidies.
Chez le moustique, le flagelle est arborescent et les scolopidies forment un double anneau entourant sa base (figure 4).
Son fonctionnement est simple. Lorsque le flagelle est sollicité par une vibration de l’air, l’action engendre un effet mécanique à sa base qui se répercute sur les scolopidies par une modification structurelle produisant finalement un potentiel d’action (PA).
Ces organes sont notamment bien développés chez les mouches (Diptères), pouvant ainsi contenir plus de 20 000 scolopides.
Les fonctions des organes de Johnston sont multiples selon les espèces et leur mode de vie :
- Indicateur de vitesse : lorsque l’insecte vole, la pression de l’air agit sur les antennes et les flagelles . Plus la vitesse est importante, plus la flexion du flagelle est grande. La vitesse est ainsi déterminée par le nombre de scolopidies stimulées.
- Recherche de partenaires sexuels : chez la drosophile par exemple, les vibrations de l’air émises par les ailes du mâle lors de la parade sexuelle sont détectées par les organes de Johnston de la femelle. La sensibilité des scolopidies est alors réglée sur la fréquence de vibration du mâle : c’est un mécanisme de reconnaissance de l’espèce. Ce même principe s’applique chez le moustique qui repère ses partenaires sexuels par les vibrations émises durant le vol.
- Communication entre individus : pour informer sur une source de nourriture, les abeilles réalisent une danse indiquant les paramètres de vol pour l’atteindre. Les vibrations sonores émises par l’abeille sont détectées par ses congénères et analysées dans leur fréquence comme dans leur intensité.
- Navigation aquatique : les coléoptères de la famille des Girinidae se déplacent à la surface de l’eau. Les organes de Johnston, en détectant les ondulations émises par leurs congénères, permettent d’éviter les collisions. Ces insectes utilisent également leurs propres vibrations sur le même principe que celui de l’écholocation des chauve-souris pour éviter les obstacles.
- Détection de la gravité : le notonecte, une punaise (Hémiptères) qui vit dans l’eau, le plus souvent postée à l’envers sous la surface, s’oriente grâce à la présence d’une bulle d’air prisonnière entre sa tête et ses antennes. Lorsque l’insecte est dans sa position normale, c’est à dire à l’envers, la bulle d’air remonte vers la surface, poussant les antennes vers le haut de la tête. La détermination de l’angle des antennes par rapport à la tête renseigne l’insecte sur sa position dans l’espace.
Les sensilles simples et les sensilles campaniformes jouent également certaines de ces fonctions : lire cet article.
IV) 4. Organes tympaniques
L’organe tympanique est un organe chordotonal spécialisé dans la réception de vibration sonore, il correspond à l’organe de l’ouïe et de l’audition.
- Structure
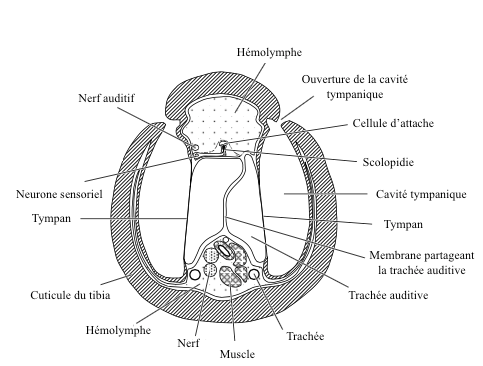
L’aspect structurel général de l’organe tympanique est simple : c’est une zone de la cuticule de faible épaisseur (le tympan ou membrane tympanique), soutenue par un sac rempli d’air (ou en contact avec une trachée) et libre de vibrer (voir figure 5 et 6).
Sur la face interne du tympan sont fixées des centaines de scolopidies qui s’activent lorsque le tympan se met à vibrer.
L’emplacement sur le corps de ces organes est très variables selon les familles et les espèces. Ils se rencontrent sur le cou de certains Coléoptères, sur la paire de pattes antérieures chez les grillons et les sauterelles de la famille des Tettigoniidae, sur l’abdomen chez les cigales (Cicadidae : lire cet article), les papillons de la famille des Geometridae et les cicindelles (Coléoptères de la famille des Cicindellidae) ou encore sur le thorax de certaines punaises aquatiques et des mantes-religieuses (Mantodea).
- Fonctionnement
Une onde sonore qui entre en contact avec le tympan le fait vibrer. Cette vibration agit sur les scolopidies, qui, selon l’amplitude et l’intensité du son, seront plus ou moins stimulés, engendrant une réponse nerveuse (PA) spécifique proportionnelle.
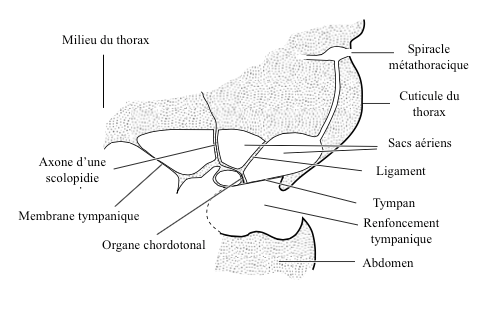
Selon la provenance, l’onde sonore peut frapper le tympan soit sur sa face externe ou interne (via les trachées ou à travers les organes), soit sur les deux faces. Ce phénomène intervient, parmi d’autres, dans la détermination de la direction de la source sonore.
La structure du tympan et la configuration de l’organe tympanique varient selon les espèces et de leur rôle. Certaines zones du tympan peuvent amplifier ou atténuer des fréquences particulières, à la manière d’un filtre acoustique. De plus, certaines scolopidies présentent des seuils d’excitabilité (de stimulation) spécifiques. Les organes tympaniques sont ainsi adaptés aux besoins des insectes.
Par exemple, la noctuelle, un papillon de la famille des Noctuelidae, possède des scolopidies sensibles à des ondes sonores de basse fréquence et d’autres à des ondes de hautes fréquence. Cette configuration lui permet de discriminer des sons d’origine lointaine (basse fréquence) correspondant au bruit environnant de ceux émis par les chauve-souris (haute fréquence) à proximité. La noctuelle peut détecter des sons d’une gamme allant de 1kHz à 140kHz, avec une sensibilité maximale autour de 20-40kHz (la fréquence moyenne émise par les chauve-souris) (voir vidéos en fin de page).
Chez d’autres insectes comme la cigale, les mâles émettent des pulsations sonores successives spécifiques à l’espèce. Les neurones des femelles sont conçus pour n’être stimulés et activés que par les fréquences et les intensités sonores propres à son espèce.
Comportement d’évitement de la noctuelle face à l’attaque d’une chauve-souris
Source :
– The Insects : Structure & Function (5ème édition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
– Keil T.A. (1997) : Functional morphology of insect mechanoreceptors – Microscopy Research and Technique, 39:506-531 (lien)
– Yack J.E. (2004) : The structure and function of auditory chordotonal organs in insects – Microscopy Research and Technique, 63:315-337 (lien)
– Encyclopedia of Insects : Vincent H. RESH & Ring T. CARDE, Academic Press-2009
– Interactions Insectes-Plantes : N. Sauvion, P.A. Calatayud, D. Thiery et F. Marion-Poll – Editions Quae : 2013
Recommandation d’ouvrages sur cette thématique
– Evolution of the Insects![]() (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
(David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
– The Insects: An Outline of Entomology![]() (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
(P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
– The Insects: Structure and Function![]() (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
(R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
– Encyclopedia of Insects![]() (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
(Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
– Interactions insectes-plantes![]() (N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae : 2013)
(N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae : 2013)
<img src= »http://wms-eu.amazon-adsystem.com/20070822/FR/img/noscript.gif?tag=passioentomo-21&linkId=3LHYKSRXQKZ3KY5I » alt= » » /><br />




Salut,
Chouette site, ça fait très plaisir de voir ça. Je signal quand même un petite erreur.
« Navigation aquatique : les coléoptères de la famille des Girinidae se déplacent à la surface de l’eau. Les organes de Johnston, en détectant les ondulations émises par leurs congénères, permettent d’éviter les collisions. Ces insectes utilisent également leurs propres vibrations sur le même principe que celui de l’écholocation des chauve-souris pour éviter les obstacles. »
L’echolocation chez le gyrin n’est qu’une hypothèse qui n’a encore jamais été réellement démontrée. Plus d’explications sont a trouver dans la thèse de Jonathan Voise : « Phénomènes capillaires et vie à l’interface air-eau. 2011. Thèse de doctorat. Tours »
Ludovic
Bonjour,
merci pour toutes ces informations très intéressantes
Benoît