Les mécanismes à l’origine de la pulvérisation défensive chez les coléoptères bombardiers
Menacés, les coléoptères bombardiers pulvérisent de leur abdomen un liquide défensif irritant par une succession de jets très rapides. Ce phénomène prend naissance dans les glandes pygidiales, où deux réactifs se mélangent, provoquant une explosion sous forte pression et la libération de ce liquide. Les mécanismes internes et anatomiques permettant cette prouesse viennent d’être décrits par l’équipe américaine d’Eric M. Arndt du Massachusetts Institute of Technology (MIT). Le contrôle des jets se fait par un mécanisme de rétroaction passif autonome ne nécessitant pas de voie nerveuse et musculaire.
Les coléoptères bombardiers appartiennent à la famille des carabes (Carabidae) et à la tribu des Barchinini (figure 1A). Ces insectes ont la faculté d’expulser un liquide défensif sous pression par des jets successifs afin d’en pulvériser sur ce qui les menace, un prédateur par exemple (voir video en fin d’article). Ce liquide défensif, issu des glandes pygidiales, est composé d’un mélange de plusieurs molécules dont les p-benzoquinones : un composé irritant employé par l’ensemble des arthropodes.
Cependant, les bombardiers sont les seuls insectes qui utilisent une réaction chimique interne explosive pour expulser cette molécule. La réaction induit également de la chaleur.
La pulvérisation du liquide est particulièrement impressionnante : une température proche de 100°C, des portées de plusieurs centimètres, une vitesse avoisinant les 10m par seconde!
Chez l’espèce Stenaptinus insignis, la fréquence d’émission des jets se situe entre 368Hz et 735Hz, donnant ainsi l’impression d’une pulvérisation unique (fréquence proche du battement des ailes d’un insecte en vol : lire cet article).
Description anatomique du système défensif
Les glandes pygidiales sont composées de trois éléments : 1) la chambre réservoir (RSC), 2) la chambre de réaction (RXC), 3) un canal de sortie (EC) donnant sur l’extrémité de l’abdomen (voir figure 1B).
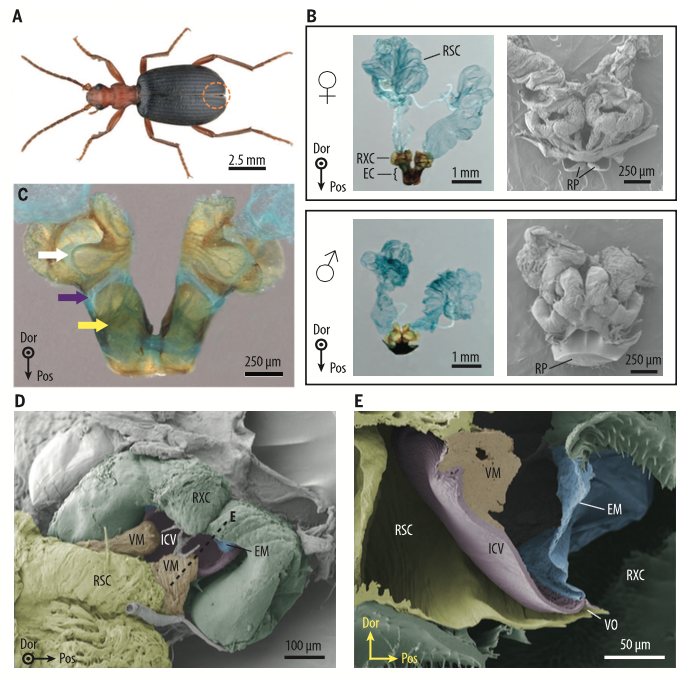
A l’extrémité du canal de sortie se trouvent des plaques réfléchissantes (RP) dont le rôle est d’orienter la direction du jet (voir figure 1B). Les deux chambres sont séparées, ainsi que leurs réactifs respectifs, par une valve dite inter-chambre (ICV) (voir figure 1D et E).
Les glandes pygidiales sont constituées de chitine (élément de la cuticule, pour en savoir plus : lire cet article) protégeant les organes internes de l’insecte des molécules chimiques toxiques, des températures élevées et des fortes pressions.
La chambre réservoir contient une solution aqueuse composée de 25% de peroxyde d’hydrogène, de 10% de p-hydroquinones et de 10% d’alcanes (non réactifs). Lorsque la valve inter-chambre s’ouvre, les réactifs de la chambre réservoir entrent en contact avec ceux de la chambre de réaction, des enzymes (peroxydase et alcalase), aboutissant à la formation des p-benzoquinones, d’oxygène gazeux, de vapeur d’eau et de chaleur.
Déroulement de la pulvérisation défensive
Eric M. Arndt et ses collègues ont pu, grâce à la microscopie optique et la numérisation électronique, décrire avec précision les structures internes et les différentes étapes du processus.
La microscopie optique a révélé une sclérotisation cuticulaire hétérogène de la chambre de réaction. Des régions sont ainsi plus flexibles ou plus rigides que d’autres. Sur la figure 1C, les zones peu sclérosées et flexibles sont en bleu, en marron celles fortement sclérosées et rigides.
Chaque émission d’une goutte de réactif de la chambre réservoir dans la chambre de réaction (ouverture de la valve inter-chambre) entraine une explosion d’une durée de 0,1ms qui génère une pression de 20kPa dans la chambre de réaction et une force de traction sur les parois de 1MPa (à titre de comparaison, des pneus de voiture sont en général gonflés à 2,2Pa).
Le contrôle des jets s’effectue au niveau de la valve inter-chambre par un phénomène d’ouverture/fermeture. Contrairement aux autres mécanismes cycliques comme celui du va-et-vient des ailes lors du vol de l’insecte (contractions alternatives d’ensemble de muscles : lire cet article sur le vol des insectes), seule l’ouverture implique l’action musculaire, la fermeture est passive : rétro-action mécanique.
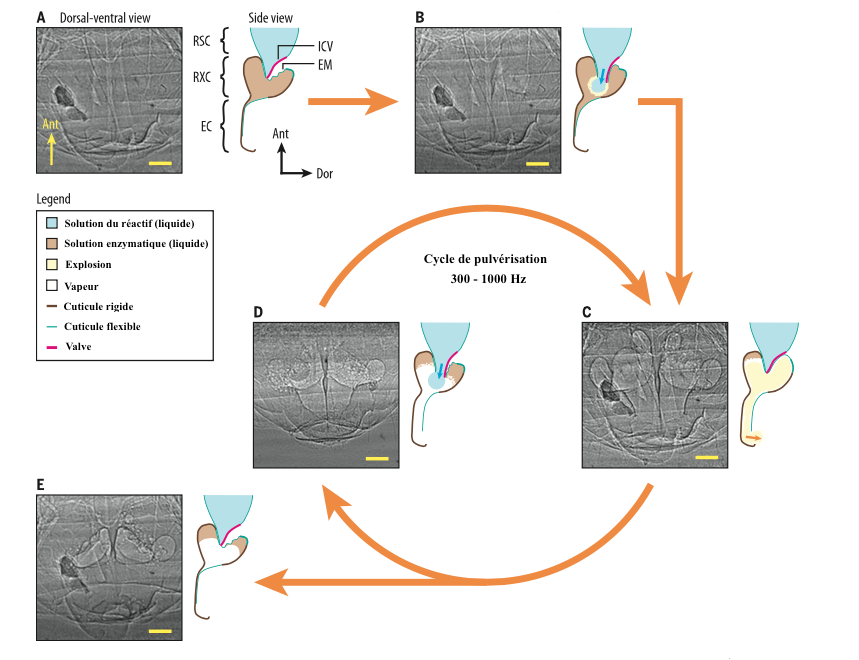
La pression dégagée par l’explosion ferme la valve inter-chambre, interrompant l’arrivée de réactif provenant de la chambre réservoir. La libération du réactif et la diminution de la pression permettent l’ouverture et l’arrivée d’une nouvelle goutte de réactif, enclenchant un nouveau cycle (voir figure 2).
La pulvérisation perdure tant que la chambre réservoir contient du réactif, ou tant que l’insecte maintient la valve inter-chambre ouverte (période de stress).
Les mécanismes internes et l’anatomie du bombardier sont remarquablement efficaces. Le contrôle des jets se fait de manière passive, sans action musculaire, par un processus de rétro-action. Ce système permet une régulation automatique des jets et de l’utilisation du réactif.
Les autres espèces proches des Barchinini émettent également des pulvérisations lors d’une menace, mais de manière continue. Les changements évolutifs à l’origine de l’apparition d’une pulvérisation saccadée, survenus chez les ancêtres des bombardiers, sont mineurs et les modifications avant tout structurelles : au niveau de la valve inter-chambre, elles n’impliquent pas d’adaptation anatomique et musculaire particulière.
Ces travaux ont été réalisés dans l’optique de concevoir de nouvelles technologies telle l’atténuation des explosions et la propulsion.
Vidéo d’une pulvérisation défensive d’un coléoptère bombardier :
Source :
– Arndt E.M. ; Moore W. ; Lee W.K. & Ortiz C. (2015) : Mechanistic origins of bombardier beetle (Brachinini) explosion-induced defensive spray pulsation. Science, vol348(6234):563-567 (lien)