
Les mouches de la famille des Diopsidae ont la particularité étonnante d’avoir les yeux positionnés à l’extrémité d’excroissances appelées pédoncules oculaires situés de chaque côté de la tête.
Cette morphologie d’hypercéphalisation et de latéralisation du système visuel se rencontre également chez des vertébrés, où le cas le plus connu est celui du requin marteau, chez des crustacés (crabes, crevettes), chez d’autres insectes comme des Hyménoptères (guêpes, abeilles), des Hétéroptères (punaises) ou encore chez d’autres familles de Diptères (mouches) : Micropezidae, Otitidae, Platystommatidae, Tephritidae, Richardiidae, Perisscelididae et Drosophilidae (voir album photos en bas de page).
La famille des Diopsidae présente toutefois le plus haut degré de complexité de ce processus adaptatif.
I) Ecologie et biologie des Diopsidae

Les Diopsidae regroupent 194 espèces réparties en 14 genres dont l’aire de répartition se concentre quasi-exclusivement sous les tropiques et l’Ancien-monde. En 1997, une espèce a été découverte en Europe : Sphyracephala europaea (voir photo 2).
Ces insectes se rencontrent dans les habitats denses et sombres des forêts tropicales et sub-tropicales, le plus souvent près de la litière. Les adultes s’alimentent de bactéries à la surface de feuilles mortes ou d’animaux morts et les larves, quant à elles, se nourrissent de débris végétaux.

Ces insectes mesurent en moyenne 10 mm de longueur. Certaines de ces espèces, comme Cyrtodiopsis whitei, ont une longueur d’écartement de leurs yeux supérieure à celle de leur corps ! (voir photos 1 et 3).
Des espèces sont dites monomorphiques (mâles et femelles possèdent des pédoncules oculaires de même longueur) et d’autres dimorphiques (les mâles ont des pédoncules oculaires plus longs que ceux des femelles). Ce dimorphisme sexuel se retrouve chez des genres et des espèces non apparentés, ce qui suggère que ce caractère morphologique serait apparu plusieurs fois indépendamment au cours de l’évolution.
Les espèces dimorphiques vivent en majorité dans la litière humide des sous-bois. Dispersées dans la végétation basse le jour, les mouches se regroupent la nuit sur le feuillage (lek). Chez des espèces comme Teleopsis dalmanni et C. whitei, à fort dimorphisme sexuel, les mâles combattent pour obtenir les meilleurs emplacements à la tombée de la nuit pour prendre le contrôle des femelles. Ces combats consistent, après observation mutuelle, à se cabrer, à déployer leurs pattes antérieures le long de leurs yeux et à les frapper (voir vidéo ci-dessous et album photos en bas de page). Ces comportements peuvent parfois se dérouler jusqu’à l’aube.

Les mouches de cette famille, contrairement à beaucoup d’autres, se reproduisent plusieurs fois tout au long de leur vie. Les femelles de T. dalmanni, par exemple, s’accouplent plus de 10 fois en une matinée. Ce comportement surprenant demeure en partie inexpliqué.
Une hypothèse suggère que les femelles multiplient les actes d’accouplement avec plusieurs mâles afin d’obtenir une quantité suffisante de sperme. Ce comportement amène par conséquent ces derniers à multiplier leurs accouplements afin d’augmenter leur chance de fertiliser une femelle, d’autant plus que les femelles de certaines espèces, comme Diasemopsis meigenii, ont la capacité d’éjecter le sperme des mâles indésirables.
De leur côté, les mâles produisent peu de spermatozoïdes, coûteux en énergie, pour pouvoir multiplier les accouplements, incitant les femelles à s’accoupler avec nombre d’entre eux. Au cours de la reproduction, les intérêts des femelles et des mâles sont de ce fait divergents : ce processus appelé « conflit sexuel », se retrouve chez tous les organismes sexués et est l’un des mécanismes à l’origine de la sélection sexuelle.
Autre particularité : le développement des Diopsidae est plus lent que pour d’autres familles de mouches, cependant, leur espérance de vie est plus longue (voir paragraphe correspondant).
II) Origine évolutive des pédoncules
La morphologie singulière de ces mouches et leur évolution ont longtemps intrigué les entomologistes :
- « Pourquoi la sélection agit-elle sur les yeux et non sur une autre partie du corps ? »

Il n’y a pas de réponse à cette question. Cependant, chez beaucoup de familles de Diptères, comme les Syrphidae, les yeux des mâles sont plus écartés que ceux des femelles. Cette caractéristique suggère à la base un rôle fonctionnel sélectionné au cours de l’évolution. Les mouches Diopsidae, comme celles de la famille des Richardiidae (voir photo 4), pousse se phénomène à l’extrême. Il est donc possible qu’une pression, un facteur écologique spécifique à ces insectes, conduise la sélection naturelle en faveur d’un déplacement des yeux à l’extrémité de pédoncules oculaires.
- « Quels sont les mécanismes évolutifs et de sélection à l’origine de tels organes ? »
Plusieurs hypothèses sont proposées :
- Hypothèse 1 : certains entomologistes, comme Burkhardt & Motte (1987), expliquent que les pédoncules oculaires apporteraient un bénéfice par une vision stéréoscopique sur une plus longue distance, 800 mm par exemple chez les mâles Cyrtodiopsis whitei, une distance plusieurs fois supérieure à celle d’autres espèces de mouches sans pédoncules comme le genre Calliphora. Cette vision à haute résolution et à large champ aurait été avantagée dans les forêts tropicales et sub-tropicales pour la recherche de congénères, rares et éloignés les uns des autres, dans un environnement sombre et dense ou pour la détection de prédateurs.
- Hypothèse 2 : l’allongement des pédoncules oculaires serait soumis à la pression de la sélection sexuelle : les femelles sélectionneraient les mâles aux pédoncules oculaires les plus longs, leur conférant un avantage sélectif. Par exemple, l’écartement des yeux serait un caractère héritable rendant les mâles sous conditions-dépendance. En d’autres termes, la longueur des pédoncules oculaires des mâles renseignerait les femelles sur la « qualité » de leur génome (ensemble des gènes), les femelles choisiraient ainsi les mâles les plus vigoureux, ceux dont le génome exprime de longs pédoncules oculaires, même lorsque les ressources alimentaires sont limitées ou peu disponibles.
- Hypothèse 3 : la fertilité des mâles serait corrélée à la longueur des pédoncules oculaires: ainsi, les femelles en choisissant les mâles aux plus longs pédoncules oculaires augmenteraient leur fitness (succès reproducteur)
Or la présence d’espèces phylogénétiquement proches monomorphiques et dimorphiques démontre l’existence de pressions de sélection autres que celle de la sélection sexuelle.
III) Système visuel
Bien que l’apparition des premiers Diptères remonterait à plus de 200 millions d’années, l’anatomie et le fonctionnement du système visuel entre les différentes familles de mouches sont restés similaires, même entre espèces éloignées phylogénétiquement.
Oeil
Contrairement à ce que l’on pourrait croire en observant l’anatomie des Diopsidae, les organes composant l’oeil : la rétine, la lame basale (lamina) et le bulbe de l’oeil composé (voir encadré et photo 5) et la structure et l’organisation des photorécepteurs (cellules sensibles aux photons), sont identiques à ceux des autres familles de mouches.
Les yeux composés (lire cet article) des mouches Diopsidae sont toutefois constitués d’un plus grand nombre d’ommatidies que la plupart des autres familles : 2 600 chez les mâles et 2 500 chez les femelles de C. whitei par exemple, et ils possèdent également une densité en photorécepteurs plus élevée.
Cette particularité offre à ces mouche une bonne acuité visuelle impliquant des adaptations anatomiques et fonctionnelles spécifiques au niveau du système nerveux central (SNC).
|
Yeux chez les insectes  Les yeux des insectes sont qualifiés de « composés » car ils sont constitués de plusieurs sous unités sensibles à la lumière appelées ommatidies : des yeux élémentaires disposés les uns à côtés des autres. Le nombre d’ommatidies par oeil est fortement variable entre les espèces, de moins d’une dizaine à plus de 20 000 chez certains Coléoptères ! L’anatomie et le fonctionnement des yeux composés ont fait l’objet de plusieurs articles : Anatomie et structure – Forme & Mouvement – Récepteurs & Polarisation |
Système nerveux central
Lorsque les cellules photoréceptrices des ommatidies sont stimulées, un influx (message) nerveux est transmis au système nerveux central (cerveau) via des axones neuronaux (nerf optique) qui relient les deux organes (lire cet article). Chez les mouches en général, un de ces axones est connecté et transmet l’information nerveuse provenant de 16 ommatidies.
Chez les Diopsidae, deux adaptations particulières sont apparues au cours de l’évolution suite à l’allongement des pédoncules oculaires. En effet, le volume disponible pour le passage de ces axones est limitée.

L’écart séparant les yeux, couplé à un nombre important d’ommatidies et une densité élevée en photorécepteurs, engendre la naissance de grandes quantités d’informations à transmettre au cerveau lorsque l’insecte se déplace (voir photo 6). Pour maintenir la rapidité de transmission de ces messages nerveux, la sélection naturelle a favorisé l’émergence d’axones à grand diamètre, diminuant d’autant l’espace disponible dans les pédoncules oculaires. Pour répondre à cette seconde contrainte, l’évolution a multiplié les connexions entre les ommatidies et les axones : ainsi, chaque axone est relié à 64 ommatidies.
Une adaptation physiologique consiste à envelopper ces axones par une gaine périneurale et par des sacs aériens afin d’optimiser l’efficacité des cellules nerveuses en fournissant davantage d’oxygène et d’éléments nutritifs. Cette adaptation se retrouve également chez tous les mammifères.
IV) Vol
L’observation du vol de certaines espèces, comme Cyrtodiopsis quinqueguttata, a montré une relative lenteur par rapport à des espèces d’autres familles, reflétant peut-être certaines limites du système nerveux ou bien dans le contrôle de l’aérodynamisme (lire cet article).
Malgré une bonne acuité visuelle et une bonne transmission de l’information nerveuse, la morphologie singulière de leur capsule céphalique entraine de nombreuses contraintes comme :
- Une vulnérabilité face aux prédateurs, à des malformations, à des accidents ou à des dommages corporels, durant le processus d’étirement et de séchage de la cuticule des pédoncules, processus de 30 min (lire cet article) (voir vidéo en bas de page)
- Une augmentation de la trainée aérodynamique et de l’instabilité lors du vol, diminuant ainsi la capacité de l’insecte à circuler dans des milieux denses et étroits
- Un fonctionnement du système visuel énergiquement coûteux en oxygène et en éléments nutritifs
- Un allongement des fibres nerveuses (axones) dans les pédoncules oculaires augmentant le temps de transmission de l’information nerveuse, ralentissant son traitement nécessaire au contrôle du vol
Une équipe américaine menée par J. F. Husak (2011, 2013) a réalisé des études du vol chez les Diopsidae. Contre toute attente, les mâles des espèces dimorphiques, comme C. whitei, volent mieux que les femelles !
Les scientifiques ont ainsi découvert un mécanisme de co-évolution compensatoire de l’allongement des pédoncules oculaires. Les mâles sont dotés d’ailes plus grandes, d’un thorax et de muscles de vol plus développés (lire cet article). Ces adaptations sont cependant absentes chez les espèces monomorphiques comme C. quinqueguttata. La corrélation existante entre la longueur des pédoncules oculaires et le développement des organes de vol indique que la sélection naturelle agit également sur tout un ensemble de gènes.
Ces scientifiques ont également mis en évidence une spécificité phylogénétique de la forme de l’aile. Bien que le phénomène d’augmentation de la taille des ailes s’est produit chez tous les genres, la forme de celles-ci a subit des voies adaptatives propres à chacun des genres, par exemple (voir figure 1 et 2) :
- Teleopsis et Diasemopsis : ailes longues et étroites
- Diasemopsis : partie distale rétrécie et bord de fuite prolongé
- Teleopsis : partie proximale rétrécie et bord distal allongé
- Sphyracephala : modèle différent, l’aile est plus large sur une grande partie de la longueur
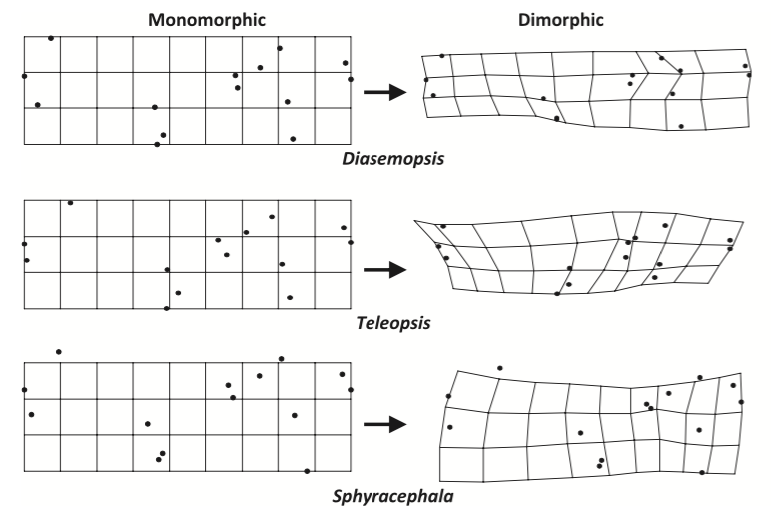
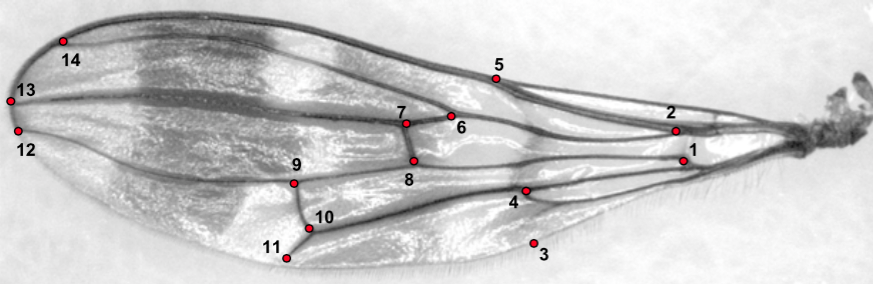
La modification de la forme de l’aile a entrainé un déplacement des nervures (lire cet article), leur épaississement à certains endroits, mais aussi un changement dans la position des cellules alaires en réponse aux contraintes aérodynamiques pour renforcer et rigidifier l’organe afin d’éviter son repliement lors du vol (voir figure 1 et 2).
V) Développement embryonnaire
Pour comprendre la morphogenèse (mise en place des différents organes) des pédoncules oculaires au stade embryonnaire chez les Diopsidae, une équipe américaine, menée par Elke E. Buschbeck (2001), a suivi le développement de la larve de Cyrtodiopsis whitei et l’a comparé à celui de Drosophila melanogaster. La drosophile est l’espèce de mouche le plus étudiée au monde, tant anatomiquement que génétiquement : elle est proche phylogénétiquement des Diopsidae.
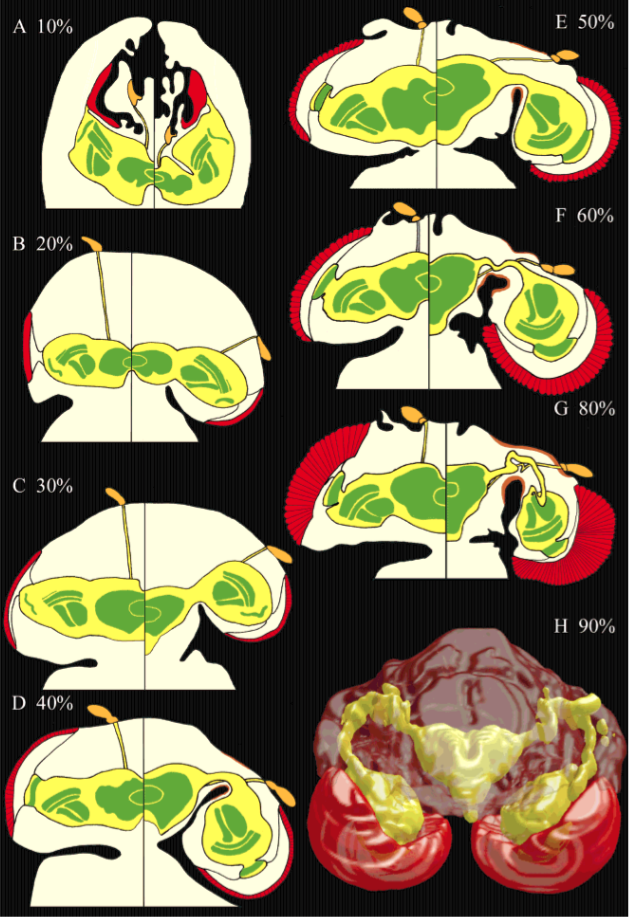
Les scientifiques ont mis en évidence l’existence de différences principalement d’ordre chronologique. Les organes du système visuel des Diopsidae apparaissent plus tôt durant le développement que chez la drosophile (voir figure 3 ci-contre).
Les lobes optiques plus imposants chez C. whitei en raison d’un nombre plus élevé d’ommatidies (2 500 contre 750 chez D. melanogaster) ne peuvent croître latéralement comme chez la drosophile mais s’orientent vers l’arrière le long de la tête.
Simultanément au déplacement des lobes optiques vers l’arrière, les pédoncules oculaires, les nerfs optiques et la cuticule s’amincissent et s’allongent, mais par manque de place, ces organes se plient et ondulent.
Au final, le développement embryonnaire des Diopsidae est deux fois plus long que celui de la drosophile en raison de la croissance des pédoncules oculaires. Il a en effet été montré que les individus de C. whitei aux pédoncules les plus courts avaient une croissance plus rapide que leurs congénères.
A l’émergence, l’allongement des pédoncules oculaires ne se fait pas par multiplication cellulaire mais uniquement par l’extension des cellules en place. Un processus qui soulève une interrogation : Quels sont les mécanismes anatomiques et génétiques qui prédifinissent la longueur finale des pédoncules oculaires ? Bien que le principe d’allongement de la cuticule et d’organes par extension soit courant chez les insectes, il est spectaculaire chez les mouches Diopsidae. Lorsque l’insecte émerge de sa pupe (chrysalide), il absorbe de l’air qu’il achemine dans les pédoncules oculaires, qui, sous pression, s’étirent tels des ballons de baudruche. Lors de cet étirement, les lobes oculaires réalisent une rotation dors-ventrale de 45°, pouvant occasionner des problèmes de malformations divers et rendant la mouche complètement vulnérable aux aléas. L’insecte reste ainsi immobile durant toute la période (30 min) du processus de rotation et de solidification de la cuticule (voir vidéo).
Le cerveau, quant à lui, est structurellement identique entre les deux espèces.
Source :
– Buschbeck E. K. & Hoy R. R. (1998) : Visual system of Stalk-Eyed fly, Cyrtodiopsis quinqueguttata : an anatomical investigation of unusual eyes – Journal of Neurobiology, p449-468 (lien)
– Buschbeck E. K. ; Roosevelt J. L. & Hoy R. R. (2001) : Eye stalk or no eye stalk : a structural comparison of pupal development in the stalk-eyed fly Cyrtodiopsis and Drosophila – The Journal of Comparative Neurology, 433:486-498 (lien)
– Chapman T. ; Pomiankowski A. & Fowler K. : Stalk-eyed flies – Current Biology, vol 15 n°14
– Husak J. F. et al. (2011) : Compensation for exaggerated eye stalks in stalk-eyed flies (Diopsidae) – Functional Ecology, 25:608-616 (lien)
– Husak J. F. et al. (2013) : Effects of ornamentation and phylogeny on the evolution of wing shape in stalk-eyed flies (Diopsidae) – Journal of Evolutionary Biology, 26:1281-1293 (lien)
– Cotton A. et al. (2014) : Male mate preference for female eyespan and fecundity in the stalk-eyed fly, Teleopsis dalmanni – Behavioral Ecology (lien)


