
Chez les insectes, les ailes sont soumises à des fortes pressions de sélection naturelle pour le camouflage, le mimétisme et la coloration d’avertissement. Les motifs et les couleurs des ailes de papillons constituent par exemple de bons modèles d’études de ces processus. Les changements au niveau génétique engendrent des modifications dans la combinaison des pigments et dans celle de leur position relative au cours du développement des ailes en phase larvaire et durant la métamorphose.
En Amérique Latine et du Sud, les papillons du genre Heliconius sont particulièrement intéressants à ce sujet. Ils présentent une large diversité de pattern (type de motif) mais qui converge dans chaque région géographique au point que toutes les espèces se ressemblent. Depuis de nombreuses années, les scientifiques essaient de comprendre quels sont les mécanismes génétiques et évolutifs à l’origine de l’apparition de cette diversité morphologique.
L’équipe, menée par Richard W.R. Wallbank (université de Cambridge), apporte des éléments de réponse. Par des analyses génétiques complexes de plusieurs espèces d’Heliconius, ces scientifiques ont mis en évidence des phénomènes de recombinaison et d’introgression de certains gènes survenus entre les espèces et les populations au cours de leur histoire évolutive. Ces mécanismes favorisent l’émergence de nouveaux patterns qui seront sélectionnés dans les populations et qui permettront ainsi aux espèces d’évoluer rapidement.
L’un des principaux obstacles à l’innovation de l’évolution est la contrainte des changements génétiques imposée par la fonction existante. En effet, les mutations conférant des phénotypes avantageux (aspect d’un trait morphologique) pour un caractère nouveau entraine souvent des effets négatifs sur d’autres traits influencés par le gène concerné (effet pléïotropique).
Ce fonctionnement pléïotropique de certains gènes restreint l’apparition de nouveautés phénotypiques et donc la diversité des espèces. Par exemple, les gènes contrôlant le développement des ailes chez les insectes ont peu été modifiés au cours de l’évolution car ils sont soumis à une forte pression de sélection. Les profils d’expression de ces gènes sont ainsi similaires entre les mouches et les papillons.

Cependant, des études ont montré qu’il existait des structures génétiques permettant de contourner ces contraintes. Elles sont constitués d’éléments juxtaposés, dits modules, codant pour des domaines spécifiques, où la modification de l’un d’entre eux ne perturbe pas l’expression des autres. Ce principe semble être à l’origine de la diversité morphologique.
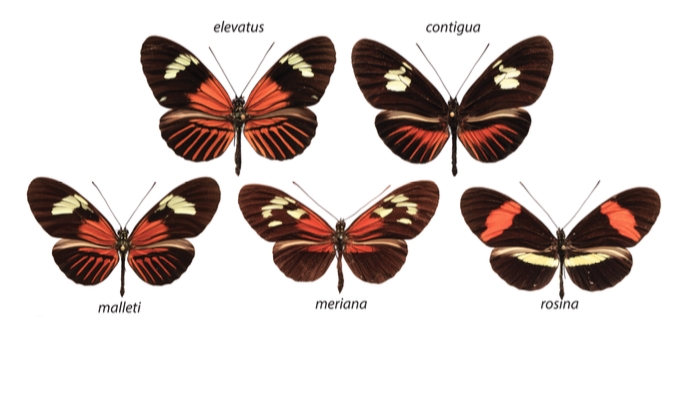
De tels éléments ont été découverts chez les Heliconius (thématique sur laquelle j’ai travaillé en 2009, rendez-vous en bas de page). Ces papillons, qui se rencontrent en Amérique Latine et du Sud, regroupent de nombreuses espèces aux motifs et aux couleurs d’une grande diversité. Dans une même localité géographique, l’ensemble des espèces présente une structuration de la coloration des ailes identique. Cette convergence mimétique est particulièrement remarquable pour les 11 espèces d’Heliconius au phénotype appelé « Dennis-Ray » : taches rouges à la base des ailes antérieures (Dennis) et rayures rouges sur les ailes postérieures (Ray) (voir figure 1).
Quels sont les processus génétiques et évolutifs qui ont ainsi permis à toutes ces espèces, parfois phylogénétiquement éloignées, d’évoluer vers un pattern similaire ?
C’est ce à quoi a voulu répondre l’équipe de Wallbank.
Pour cela, les scientifiques ont entrepris des analyses génétiques sur Heliconius melpomene et d’espèces proches comme H. elevatus et H. timareta pour identifier les modules associés aux éléments à l’origine des phénotypes Dennis et Ray.
Leurs résultats révèlent une histoire évolutive plus complexe que les scientifiques le supposaient !
Les analyses génomiques ont été réalisées à partir de 96 spécimens appartenant à une population de H. melpomene provenant de Guyane : H. m. meriana qui ne présente pas de rayures sur les ailes postérieures ; à une population de H. timareta provenant d’Equateur : H. t. timareta forme contigua n’ayant pas de motifs Dennis sur les ailes antérieures ; à des individus hybrides de H. melpomene d’Equateur ne possédant pas de rayures sur les ailes postérieures (voir figure 2).
A l’issu de ces analyses, les scientifiques ont découvert deux régions distinctes, chaque séquence étant associée à l’un des deux phénotypes (10 000 et 25 000 paires de bases pour les phénotypes Dennis et Ray respectivement) (voir figure 3).
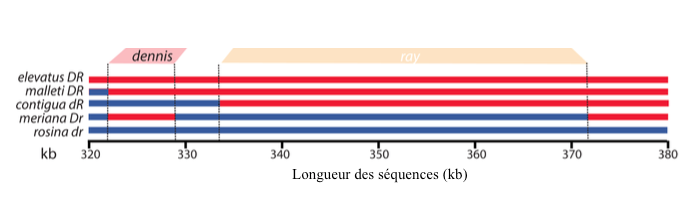
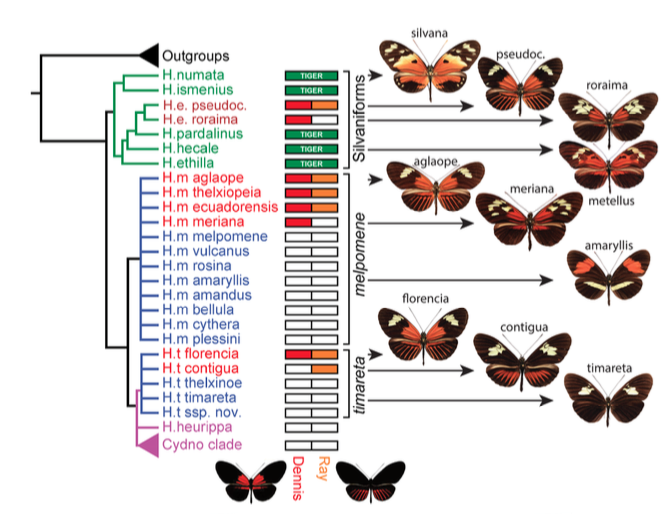
Ensuite, à l’aide d’une méthode appelée Base Local Alignment Search Tool (BLAST), les scientifiques ont aligné des différentes séquences de ces individus pour les comparer entre elles mais aussi avec celle de H. cydno, n’ayant pas de rayures et celle d’une espèce de référence, plus éloignée phylogénétiquemetn et appartenant au clade des Silvaniformes : H. elevatus.
Leurs résultats indiquent que les phénotypes Dennis et Ray sont contrôlés par des éléments génétiques adjacents distincts, où chacun d’eux s’exprime différemment au sein de chaque espèce et sous-population d’Heliconius, leur apportant ainsi un pattern spécifique.
A l’aide de ces nouvelles sur l’architecture génétique des allèles (version d’un gène) à l’origine des phénotypes Dennis et Ray, les scientifiques ont pu reconstituer l’histoire évolutive de ces éléments génétiques au sein de ces espèces. Celle-ci repose sur de multiples phénomènes successifs et récents d’introgression (échange de gène entre espèces) et de recombinaison entre les trois espèces : H. elevatus, H. melpomene et H. timareta (voir figure 4).
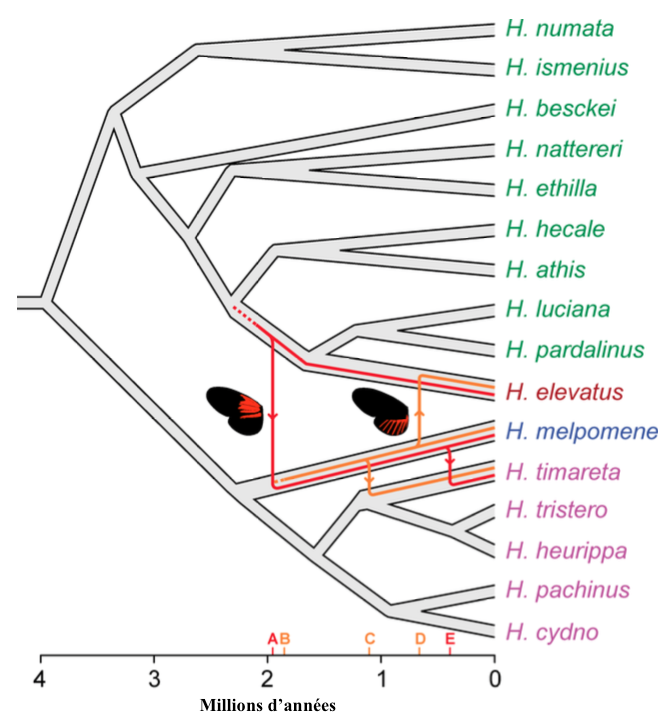
L’allèle Dennis actuellement présent chez H. melpomene proviendrait d’une forme ancestrale de H. elevatus, une transmission survenue il y a 2 millions d’années, soit 2 millions d’années après que ces espèces aient divergé. Les espèces du clade Silvaniforme miment des papillons de la tribu des Ithomiini et sont d’apparence marbrée, orangée/rouge avec des motifs noirs et jaunes « tigrés ». Les tâches orangées à la base de leurs ailes antérieures, similaires aux phénotypes Dennis de H. melpomene, semblent confirmer cette origine ancestrale.
Un second événement d’introgression de cet allèle serait survenu plus récemment, 0,45 millions d’années, vers H. timareta.
L’allèle à l’organe du phénotype Ray aurait été transmis une première fois de H. melpomene vers H. timareta il y a un peu plus d’un million d’années, puis une seconde fois vers H. elevatus 400 000 ans plus tard.
Ainsi, au cours de l’évolution, les pattern alaires de ces espèces se modifient. Par exemple, la forme Dennis était la seule représentée dans les populations de Guyane durant 1 million d’années pour ensuite évoluer vers la forme actuelle Dennis-Ray.
Les faibles niveaux de diversité génétique entre ces allèles au sein des espèces confirment l’origine récente de ces introgressions : les allèles ne différent que de moins de 2% (horloge moléculaire : hypothèse selon laquelle les mutations génétiques s’accumulent dans un génome à une vitesse globalement proportionnelle au temps géologique).
Remarques : une forme de H. melpomene, H. m. meriana, ne présente plus le phénotype Ray. La perte de ce phénotype résulte d’une recombinaison au niveau de cet allèle l’empêchant de s’exprimer. Le même événement est survenu pour les allèles Dennis et Ray chez H. t. timareta, quasiment entièrement noire (voir figure 5).
Cette étude apporte de nouveaux éléments pour mieux comprendre comment l’évolution fonctionne et met en évidence les mécanismes génétiques à l’oeuvre conduisant à l’apparition de nouvelles formes morphologiques. Les introgressions et les recombinaisons semblent expliquer l’histoire évolutive d’autres espèces comme les pinsons de Darwin, des épinoches ou encore … l’homme.
Source :
– Wallbank R.W.R. et al. (2016) : Evolutionary novelty in a butterfly wing pattern through enhancer shuffling. PLoS Biol. 14(1):1-16 (lien)
Albums photos et vidéos d’Heliconius
Photos et vidéos prises par Benoît GILLES lors de ses séjours au Panama et Guyane française dans le cadre d’études sur les Heliconius au Muséum National d’Histoire Naturelle de Paris : travaux de recherche ayant fait l’objet d’une publication à lire ici


Billet tout à fait fascinant… merci!
Petite remarque « pattern » se traduit simplement par « patron », notamment « patron alaire » (c’est le terme dédié en biologie), patron de couture, etc.
Bonjour, merci pour votre compliment ! Vous avez raison concernant le mot « patron ». C’est un choix personnel d’avoir garder le mot pattern. Benoît GILLES