2020, l’invasion du Criquet pèlerin en Afrique de l’Est : récurrence d’un phénomène historique
Par Antoine Foucart ; Pierre-Emmanuel Gay & Cyril Piou
L’invasion du Criquet pèlerin en Afrique de l’Est depuis le début 2020 est préoccupante. C’est la pire depuis des décennies.
L’Organisation des Nations Unies pour l’alimentation et l’agriculture (FAO) a appelé à l’aide internationale dès janvier pour réunir 70 millions de dollars US destinés à financer hommes, matériels et opérations de lutte. Les coûts estimés par la FAO s’élèvent maintenant à plus de 300 millions de dollars US. La Banque Mondiale a également débloqué 500 millions de dollars US pour faire face à ce problème. Des dizaines de milliers d’hectares de terres cultivées et de pâturages ont déjà été ravagés en Éthiopie, en Somalie et au Kenya. Plus de 20 millions de personnes sont potentiellement en état d’insécurité alimentaire. Plus à l’Est, les essaims de criquets envahissent depuis deux ans plusieurs pays de la Péninsule Arabique, le Pakistan, l’Iran et l’Inde. Ils sont également susceptibles de progresser vers l’Ouest en suivant les vents : l’est de l’Ouganda et le Soudan du Sud sont déjà atteints. Le Tchad puis l’Afrique de l’Ouest et du Nord pourraient être menacés dans les mois qui viennent.
Qui est le Criquet pèlerin ?
Le Criquet pèlerin (Schistocerca gregaria Forsskål 1775) est un locuste, c’est-à-dire qu’en fonction des conditions environnementales qu’il rencontre, cet orthoptère a la capacité de s’adapter à son milieu en présentant deux formes morphologiques distinctes, avec des comportements opposés : la phase solitaire et la phase grégaire. Les criquets solitaires, de couleur brun sable (illustration 1A), ne cherchent pas le contact avec leurs congénères et se déplacent plutôt à la tombée de la nuit.
Les grégaires ont des couleurs vives : jaune et noir pour les larves, rose lorsqu’ils sont ailés et sexuellement immatures (illustration 1C), jaune à maturation (illustration 1B) ; ils vivent en groupes et se déplacent le jour. Jusqu’au début du XXe siècle, on pensait même que solitaires et grégaires ne faisaient pas partie de la même espèce. C’est l’entomologiste russo-anglais Sir Boris Uvarov (1886-1970) qui a découvert en 1928 la capacité de certains criquets à changer entre ces deux phases (polyphénisme de phase).

La plupart du temps, les Criquets pèlerins ne font pas parler d’eux. Ils vivent naturellement isolés les uns des autres, à l’écart des populations humaines, en très faible densité, dispersés sur les immenses surfaces désertiques et semi-désertiques de l’Afrique et de l’Asie du Sud-Ouest, depuis la Mauritanie jusqu’à la frontière indo-pakistanaise (illustration 2). En phase solitaire, ces criquets fréquentent les fonds d’oueds ou encore le pied des reliefs où ils trouvent le peu d’eau résiduelle qui fera pousser une maigre végétation qui leur suffira à se nourrir. Ils ne posent alors aucun problème et sont presque invisibles pour l’homme. Leur faible nombre ainsi que leur isolement limite leur prélèvement sur la biomasse. Les cultures vivrières des pays concernés ne sont pas touchées.
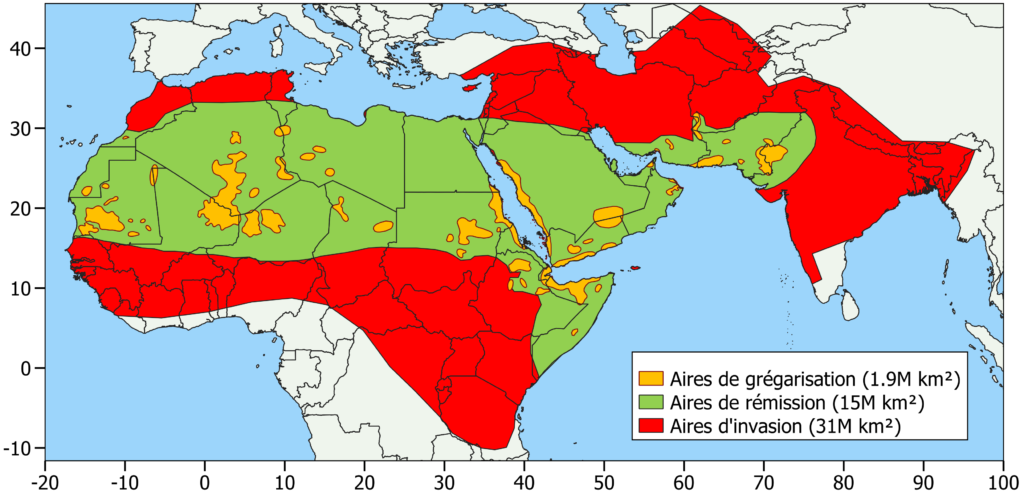
Mais lorsqu’à la suite d’événements climatiques favorables (pluies, températures, vents) des populations de criquets sont amenées à se regrouper et que leur densité atteint un seuil critique, les nombreux contacts interindividuels induisent un changement de comportement graduel, que l’on appelle la grégarisation. Les criquets grégarisant deviennent plus actifs et plusieurs processus auto-catalytiques de synchronisation et de choix de sites de pontes favorisent le renforcement des caractéristiques grégaires.
Lorsque leur densité est suffisante, les grégaires initient alors des mouvements de groupe. Les jeunes forment des bandes larvaires (illustration 3 ci-dessous), soit de gigantesques tapis mouvants comptant des milliers d’individus dépourvus d’ailes et se déplaçant de quelques centaines de mètres chaque jour. Devenus adultes, ils forment des essaims d’ailés et parcourent de longues distances (jusqu’à 150 km par jour) en suivant les vents dominants et les fronts pluvieux, comme le déplacement saisonnier du front intertropical, pour trouver de la nourriture fraîche (vidéo ci-dessous). Les essaims peuvent atteindre des dimensions de l’ordre de plusieurs dizaines de kilomètres carrés voire davantage, et compter des milliards d’individus. Ils dévastent toute la végétation, cultures vivrières comprises, sur leur passage.
On estime qu’un essaim de 1 km² compte 50 millions d’individus qui dévorent 100 t de végétation chaque jour, soit autant que la nourriture de 1500 vaches. On a vu récemment au Kenya des essaims de plusieurs centaines de kilomètres carrés. C’est donc un problème majeur de sécurité alimentaire pour les populations concernées.
La genèse de l’invasion actuelle

L’invasion actuelle résulte principalement du passage du cyclone Luban sur les côtes yéménites et omanaises en octobre 2018, qui y a déversé de fortes pluies, lesquelles ont favorisé une explosion de la végétation dans ces zones semi-désertiques. Cette nourriture abondante a attiré les criquets qui s’y sont regroupés puis reproduits de façon exponentielle. D’une génération à l’autre, soit un trimestre, leur nombre peut être multiplié par 20 dans des conditions idéales.
Dès le début 2019, de petits essaims se sont formés puis répandus au travers de la péninsule Arabique vers l’Iran, le Pakistan puis l’Inde. D’autres restés sur place ont continué à se multiplier. En juin 2019, ils se sont répandus de l’autre côté du Golfe d’Aden et de la Mer Rouge, vers la Corne de l’Afrique. Ils y ont envahi l’Éthiopie et la Somalie. Les inondations d’octobre et de novembre dans la région y ont créé de bonnes conditions pour qu’ils continuent de se multiplier. Ainsi, à partir de décembre 2019, ils ont envahi le Kenya jusqu’au nord de la Tanzanie, ainsi que l’Ouganda et le Soudan du Sud.
La récurrence des invasions
Même si de nombreuses plumes se sont attelées à relier l’invasion actuelle aux changements climatiques, les invasions de Criquets pèlerins sont un phénomène naturel documenté par l’homme depuis des millénaires : ils constituaient la 8ème plaie d’Égypte citée dans l’Ancien Testament et on les retrouve même dessinés sur des bas-reliefs des pyramides. Ces insectes ont probablement constitué un problème pour les humains sédentaires depuis les débuts de l’agriculture. Leurs invasions font régulièrement des ravages mais n’ont pas de cycle régulier : elles se produisent dès que les conditions écologiques deviennent favorables aux criquets, au premier rang desquelles la pluviométrie. Les invasions acridiennes sont donc à considérer comme une conséquence d’aléas ou de phénomènes climatiques récurrents.
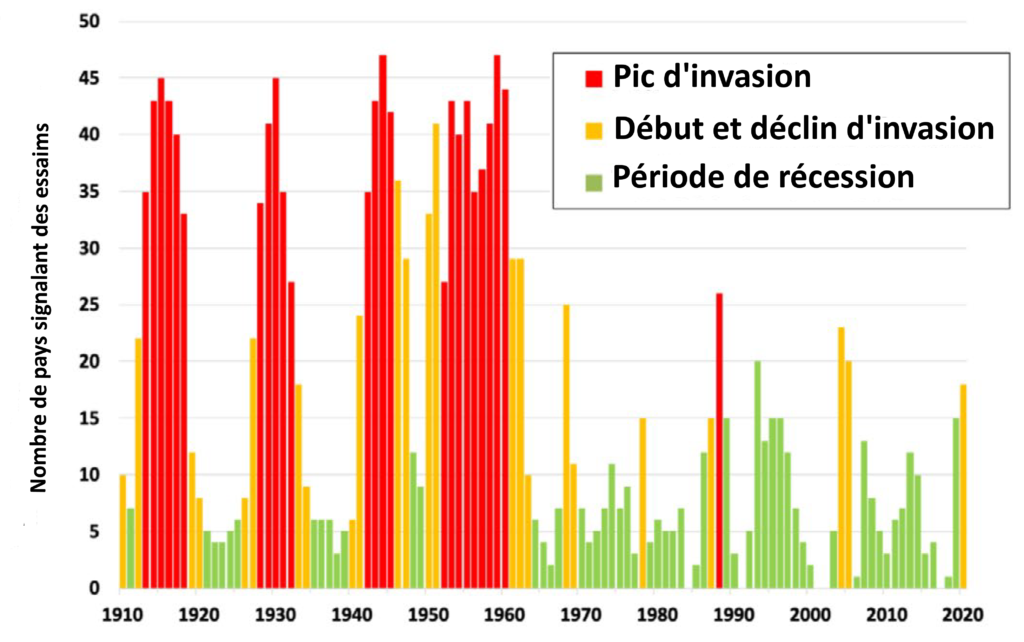
Jusqu’aux années 1950, les invasions se déroulaient et s’éteignaient parfois d’elles-mêmes sous l’effet de conditions naturelles devenues moins favorables. Mais le processus prenait plusieurs années et pouvait s’enclencher avec une telle fréquence que les cycles d’invasion (illustration 4) semblaient ne jamais finir complètement au regard de la zone de répartition de l’espèce. Certaines invasions (il y en a eu cinq majeures dans la 1ère moitié du XXe siècle) ont duré jusqu’à treize ans. Afin de protéger les cultures, les hommes ne sont parvenus à adopter des mesures de lutte active puis de gestion préventive qu’à partir des années 1960, notamment grâce à l’utilisation coordonnée au niveau international de la lutte chimique et du développement de recherches afin de mieux connaitre le Criquet pèlerin et son écologie.
Depuis, même si les invasions n’ont pas été éradiquées, elles sont moins fréquentes et surtout moins longues : il n’y en a eu ainsi que trois, toutes de moins de trois ans, dans la deuxième moitié du XXème siècle. Si ces invasions et les deux du XXIème siècle (2003-2005 et actuellement) sont le résultat de conditions climatiques favorables, elles correspondent également à des moments où la gestion préventive a été négligée, oubliée ou impossible à maintenir à cause de conflits armés dans certaines zones de l’aire de distribution du Criquet pèlerin.
La solution de la gestion préventive
Surveillance et maîtrise des populations acridiennes sont donc les deux points clés de la gestion préventive. C’est le maintien permanent des effectifs de l’insecte en-deçà du seuil de densité déclenchant la transformation phasaire qui permet d’éviter la formation d’essaims importants.
Tout le problème est de savoir où et quand celle-ci va s’effectuer. On parle ici de zones désertiques ou semi-désertiques à l’échelle continentale, peu fréquentées par l’homme donc sans témoignages directs ou alors très tardifs. Il faut aussi intervenir à temps : les larves marchent, un essaim vole. Plus on s’y prend tôt, plus on peut intervenir à petite échelle, de manière localisée pour détruire les insectes en cours de regroupement afin de casser le phénomène de grégarisation. En Mauritanie par exemple, les équipes de lutte traitent quelques centaines de km carrés chaque année depuis 2007, et ça fonctionne ! Si on attend que les larves presque statiques deviennent des essaims mobiles, c’est beaucoup plus difficile et coûteux de les réguler. Et dévastateur pour l’environnement.
Ce qui permet la gestion préventive, c’est l’addition, année après année :
- de l’expérience des prospecteurs envoyés par les centres antiacridiens de leurs pays respectifs et qui parcourent régulièrement le terrain,
- des résultats de la recherche scientifique qui ont permis de connaitre l’écologie de l’insecte et de dresser des cartes de zones favorables aux débuts d’invasion,
- de l’imagerie satellitaire qui détecte toute pluie ou humidité qui permet le développement de la végétation, laquelle assure des conditions de développement idéales à des insectes de plus en plus nombreux.
Les Nations Unies, au travers de la FAO, gèrent les actions de gestion préventive au niveau continental. Le Desert Locust Information Service (DLIS) basé à Rome, émet des prévisions pour les pays d’Afrique et d’Asie touchés par le Criquet pèlerin. Trois commissions (CLCPRO-Commission de lutte contre le Criquet pèlerin en région occidentale à Alger, CRC-Commission pour la Région centrale au Caire pour l’Afrique de l’Est et le Proche Orient, SWAC-South West Asia Commission à Rome pour l’Asie du Sud-ouest) coordonnent ensuite les actions au niveau régional. Ces commissions s’appuient sur le travail constant d’unités de lutte antiacridiennes nationales. Plus ces unités sont indépendantes et bien financées au sein des pays, plus elles arrivent à maintenir une surveillance et une réponse précoce efficaces.
Nuée de criquets migrateurs (Locusta migratoria) – Madagascar – 2016 – Benoît GILLES
Depuis 2005, c’est un succès en Afrique de l’Ouest et du Nord-ouest, où les actions sont coordonnées par la CLCPRO dans 10 pays : Algérie, Burkina Faso, Libye, Mali, Maroc, Mauritanie, Niger, Sénégal, Tchad et Tunisie. La CLCPRO a poussé ces pays à collaborer de façon permanente en échangeant des informations et en maintenant l’attention et l’intérêt, même en période de calme acridien, avec des activités de recherche et de formation.

Il en va autrement en Afrique de l’Est où la gestion préventive est assurée non pas par une mais deux institutions séparées : la Commission pour la région centrale de la FAO (CRC) et le Desert Locust Control Organization for Eastern Africa (DLCO-EA). Seuls certains pays sont membres des deux institutions. Malheureusement, la prospection préventive fait défaut dans plusieurs pays, et pas seulement ceux en instabilité politique. Des actions de formation, des réinvestissements dans des véhicules et des outils de prospection de qualité sont nécessaires.
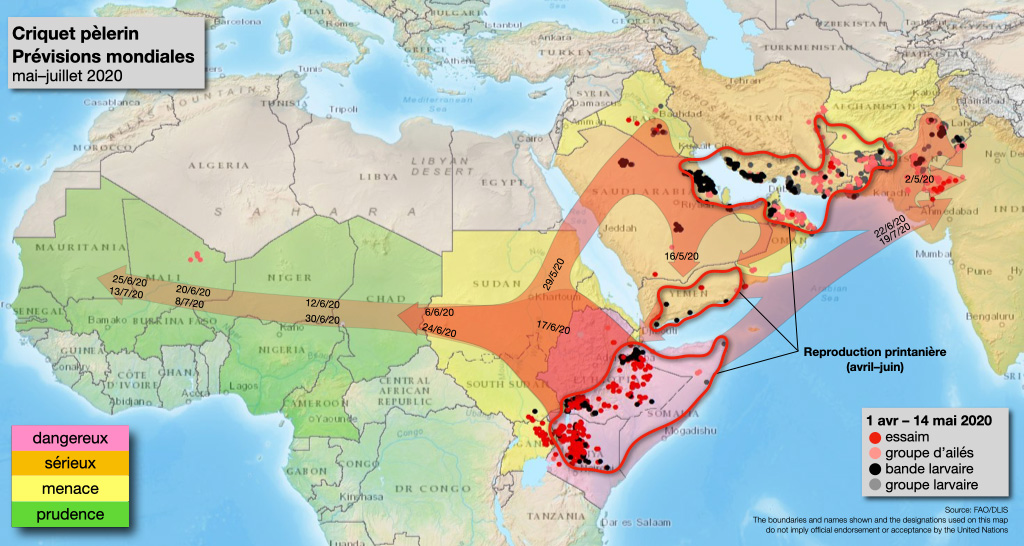
Mais c’est surtout l’instabilité sécuritaire de ces régions qui explique l’aspect catastrophique de la situation actuelle, où aucune mesure coordonnée n’a pu être prise. Les essaims ont commencé à se constituer au Yémen mais la guerre a empêché toute gestion de la crise. En arrivant en Somalie, un pays bien trop déstabilisé pour avoir une gestion préventive efficace, les criquets ont continué à se multiplier (illustration 5). Le Kenya, relativement peu fréquenté habituellement par le Criquet pèlerin s’est rapidement retrouvé submergé malgré ses structures de recherche.
La recherche pour améliorer la gestion préventive
La recherche contribue à l’amélioration du système de gestion préventive. Elle peut intervenir sur la prévision des risques avec des images satellitaires et de la modélisation (illustration 6). Elle permet aussi de mieux analyser l’écologie des acridiens. En étudiant les déplacements de groupe, en analysant leur utilisation de la végétation et en comprenant leurs comportements de thermorégulation, on peut mieux déterminer le lieu et le meilleur moment pour appliquer les pesticides, afin de provoquer un plus fort taux de mortalité et un impact environnemental plus faible.
La recherche permet également de maintenir les systèmes de prévention en état de vigilance permanente. Les crises apparaissent tous les 10-15 ans et sont liées en partie au manque de prévention. Cela arrive lorsque les activités de gestion ne sont plus médiatisées depuis plusieurs années et lorsque l’attention et l’intérêt des acteurs s’émoussent, que les financements se font plus rares, que les systèmes de gestions, moins sollicités, se mettent en léthargie, se désorganisent, etc.
Depuis les années 1970 le Centre international de recherche agronomique pour le développement (CIRAD, Montpellier, France), à la suite des instituts qui l’ont précédé, a maintenu une expertise sur l’écologie et la gestion des acridiens et en particulier sur le Criquet pèlerin. Nos travaux ces 15 dernières années, depuis la dernière invasion de 2003-2004, ont permis de repenser la gestion préventive (Lecoq 2005, 2008 ; Doré et al. 2005, 2008 ; Magor et al. 2008 ; Sword et al. 2010), de mieux intégrer les migrations de solitaires et les différences entre populations de criquets en utilisant des outils moléculaires (Chapuis et al. 2014, 2015, 2016, 2017, 2020) et de mieux comprendre le rôle de la végétation dans les déplacements de bandes larvaires (Dkhili et al. 2017, 2019).
Les coopérations avec le JIRCAS (Japanese International Research Center for Agricultural Sciences) du Japon ont permis d’étudier plus finement les comportements de terrain (Maeno et al. 2012, 2013a) et la plasticité des femelles à changer leurs allocations énergétiques dans les œufs (Maeno et al. 2013b, 2020).
Nos travaux ont aussi servi de manière plus opérationnelle à identifier les seuils de densités de grégarisation du Criquet pèlerin (Cissé et al. 2013, 2015, 2016) et à évaluer le rôle de différents facteurs environnementaux dans les dynamiques de populations afin d’aider à l’orientation des équipes de terrains (Piou et al. 2013, 2017, 2019 ; Lazar et al. 2016 ; Kayalto et al. 2020). Et finalement, différentes approches de modélisation ont permis de mieux comprendre les dynamiques de population de ce criquet (Akimenko & Piou 2018) et du système de gestion (Gay et al. 2018, 2020), de débuter des travaux sur les risques à venir liés aux changements climatiques (Meynard et al. 2017, 2020), et d’aider à la prévision des déplacements d’essaims (Piou 2020) (illustration 5).
La particularité d’une majorité de ces travaux est aussi qu’ils ont été conduits en partenariat avec les acteurs de la gestion préventive en Afrique de l’Ouest et du Nord, notamment avec la CLCPRO. Ce maintien d’une activité de recherche avec et dans les centres antiacridiens du Maroc, de Mauritanie, du Mali, du Tchad ou d’Algérie ont permis un transfert de connaissance et un maintien d’expertise malgré certaines années d’absence totale de population grégaires sur le terrain. La recherche, n’étant pas en dehors du système de gestion, a donc une responsabilité pour maintenir cette vigilance permanente.
Vidéo FAO
Conclusion
Même si les invasions du Criquet pèlerin sont si spectaculaires et importantes économiquement qu’elles mobilisent des centaines de millions de dollars tous les 10-15 ans, les acteurs de la recherche sont, entre ces crises et malgré des ressources souvent assez dérisoires en regard du risque, des membres importants du système préventif. La disparition dans les années 1990 du Centre for Overseas Pest Research britannique, héritier de l’Anti-Locust Research Center que sir Boris Uvarov a dirigé pendant de nombreuses années, a marqué un déclin dans la recherche opérationnelle avec et pour les gestionnaires.
À la même époque, le CIRAD comptait jusqu’à vingt personnes à Montpellier et des centaines de collaborateurs de par le monde travaillant sur les criquets ravageurs. Depuis, les effectifs n’ont cessé de baisser avec maintenant un collectif de dix personnes (dont seulement quatre cadres permanents). Pourtant, la littérature scientifique sur les locustes a augmenté exponentiellement mais en favorisant les aspects purement académiques et déconnectés des besoins de compréhension des gestionnaires. Il faut espérer que cette crise sans précédent permette une prise de conscience de l’importance de la recherche de qualité pour trouver de nouvelles solutions, plus respectueuses de l’environnement, dans la gestion des populations de criquets ravageurs.
Lien
Bibliographie
- Akimenko V.V. & Piou C. (2018) : Two-compartment age-structured model of solitarious and gregarious locust population dynamics. Mathematical Methods in the Applied Sciences, 41: 1-37 (lien)
- Chapuis M.P. ; Plantamp C. ; Blondin L. ; Pagès C. ; Vassal J.M. & Lecoq M. (2014). Demographic processes shaping genetic variation of the solitarious phase of the Desert locust. Molecular Ecology, 23(7): 1749-1763 (lien)
- Chapuis M.P. ; Plantamp C. ; Streiff R. ; Blondin L. & Piou C. (2015) : Microsatellite evolutionary rate and pattern in Schistocerca gregaria inferred from direct observation of germline mutations. Molecular Ecology, 24:6107-6119 (lien)
- Chapuis M.P. ; Bazelet C.S. ; Blondin L. ; Foucart A. ; Vitalis R. & Samways M.J. (2016) : Subspecific taxonomy of the Desert locust, Schistocerca gregaria (Orthoptera: Acrididae), based on molecular and morphological characters. Systematic Entomology, 41(3), 516-530 (lien)
- Chapuis M.P. ; Foucart A. ; Plantamp C. ; Leménager N. ; Benoit L. ; Gay P.E. & Bazelet C.S. (2017) : Genetic and morphological variation in non-polyphenic southern African populations of the Desert locust. African Entomology, 25(1): 13-23 (lien)
- Chapuis M.P. ; Raynal . ; Plantamp C. ; Meynard C.M. ; Blondin L. ; Marin J.M. & Estoup A. (2020) : A young age of subspecific divergence in the Desert locust Schistocerca gregaria, inferred by ABC Random Forest. bioRxiv: 671867(ver. 4/peer-reviewed and recommended by PCI Evolutionary Biology, 2020), 1-36 (lien)
- Cissé S. ; Ghaout S. ; Mazih A. ; Ould Babah M.A. ; Sidi Benahi A.S. & Piou C. (2013) : Effect of vegetation on density thresholds of Desert locust gregarization from survey data in Mauritania. Entomologia Experimentalis et Applicata, 149:156-165 (lien)
- Cissé S. ; Ghaout S. ; Mazih A. ; Jourdan-Pineau H. ; Maeno K. & Piou Cyril. (2015) : Characterizing phase-related differences in behaviour of Schistocerca gregaria with spatial distribution analysis. Entomologia Experimentalis et Applicata, 156:128-135 (lien)
- Cissé S. ; Ghaout S. ; Babah Ebbe M.A. ; Kamara S. & Piou C. (2016) : Field verification of the prediction model on Desert locust adult phase status from density and vegetation. Journal of Insect Science, 16(174):1-4 (lien)
- Dkhili J. ; Berger U. ; Idrissi Hassani M. ; Ghaout S. & Piou C. (2017) : Self-organized spatial structures of locust groups emerging from local interaction. Ecological Modelling, 361: 26-40 (lien)
- Dkhili J. ; Maeno K. ; Idrissi Hassani M. ; Ghaout S. & Piou C. (2019) : Effects of starvation and vegetation distribution on locust collective motion. Journal of Insect Behavior, 32: 207-211 (lien)
- Gay P.E. ; Lecoq M. & Piou C. (2018) : Improving preventive locust management: insights from a multi-agent model. Pest Management Science, 74(1), 46-58 (lien)
- Gay P.E. ; Lecoq M. & Piou C. (2020) : The limitations of locust preventive management faced with spatial uncertainty: exploration with a multi-agent model. Pest Management Science, 76(3): 1094-1102 (lien)
- Kayalto M. ; Idrissi Hassani M. ; Lecoq. ; Gay P.E. & Piou C. (2020) : Cartographie des zones de reproduction et de grégarisation du criquet pèlerin au Tchad. Cahiers Agricultures. 200029 : 1-10
- Lazar M. ; Piou C. ; Doumandji-Mitiche B. & Lecoq M. (2016) : Importance of solitarious Desert locust population dynamics: lessons from historical survey data in Algeria. Entomologia Experimentalis et Applicata, 161(3) : 168-180 (lien)
- Lecoq M. (2005) : Threat of bad bugs: can we avoid Desert Locust crises? Environment and Poverty Times, 03: 13 (lien)
- Lecoq M. (2008) : Stratégie et logistique de la lutte contre le Criquet pèlerin. Biotechnology for locust control, Rabat, Maroc, 1-3 juillet 2008
- Maeno K. ; Piou C. ; Ould Ely S. ; Ould Babah M.A. ; Pélissié B. ; Ould Mohamed S.A. ; Javar Bacar M.E.H. ; Etheimine M. & Nakamura S. (2013a) : Plant size-dependent escaping behaviour of gregarious nymphs of the Desert locust, Schistocerca gregaria. Journal of Insect Behavior, 26(5): 623-633 (lien)
- Maeno K. ; Piou C. ; Ould Babah M.A. & Nakamura S. (2013b) : Eggs and hatchlings variations in Desert locusts: phase related characteristics and starvation tolerance. Frontiers in Physiology, 4(345), 1-10 (lien)
- Maeno K. Piou C. & Ghaout S. (2020) : The desert locust, Schistocerca gregaria, plastically manipulates egg size by regulating both egg numbers and production rate according to population density. Journal of Insect Physiology, 122(104020): 1-10 (lien)
- Magor J.I. ; Lecoq M. & Hunter D.M. (2008) : Preventive control and Desert locust plagues. Crop Protection, 27(12): 1527-1533 (lien)
- Meynard C. ; Gay P.E. ; Lecoq M. ; Foucart A. ; Piou C. & Chapuis M.P. (2017) : Climate-driven geographic distribution of the Desert locust during recession periods: Subspecies’ niche differentiation and relative risks under scenarios of climate change. Global Change Biology, 23(11): 4739-4749 (lien)
- Meynard C.M. ; Lecoq M. ; Chapuis M.P. & Piou C. (2020) : On the relative role of climate change and management in the current desert locust outbreak in East Africa. Global Change Biology, 1-3 (lien)
- Piou C. ; Lebourgeois V. ; Sidi Benahi A.S. ; Bonnal V. ; Javar Bacar M.E.H. ; Lecoq M. & Vassal J.M. (2013) : Coupling historical prospection data and a remotely-sensed vegetation index for the preventative control of Desert locusts. Basic and Applied Ecology, 14(7): 593-604 (lien)
- Piou C. ; Jaavar Bacar M.E.H. ; Babah Ebbe M.A. ; Chihrane J. ; Ghaout S. ; Cissé S. ; Lecoq M. & Ben Halima T. (2017) : Mapping the spatiotemporal distributions of the Desert Locust in Mauritania and Morocco to improve preventive management. Basic and Applied Ecology, 25: 37-47 (lien)
- Piou C. ; Gay P.E. ; Sidi Benahi A.S. ; Ould Babah M.A. ; Chihrane J. ; Ghaout S. ; Cissé S. ; Diakité F. ; Lazar M. ; Cressman K. ; Merlin O. & Escorihuela M.J. (2019) : Soil moisture from remote sensing to forecast Desert locust presence. Journal of Applied Ecology, 56(4): 966-975 (lien)
- Piou C. (2020) : Modélisation du risque d’arrivée d’essaims au Tchad au premier semestre 2020. https://locustcirad.wordpress.com
- Sword GA.. ; Lecoq M. & Simpson S.J. (2010) : Phase polyphenism and preventative locust management. Journal of Insect Physiology, 56: 949-957 (lien)