
Les termites : Biologie
Par Guillaume BAUDOUIN
Généralités
|
L’auteur : Guillaume Baudouin Suite à un parcours universitaire entre Angers, Rennes et Poitiers, Guillaume a réalisé sa thèse sur la problématique des termites à Paris dans le cadre d’un projet mené entre l’IRBI (Institut de Recherche sur la Biologie de l’Insecte) et la Mairie de Paris entre 2013 et 2017 Interview de l’auteur en bas de page |
Les termites appartiennent au super-ordre des Dictyoptères. Ils forment l’ordre des Isoptera et sont apparentés aux Blattes. Certains auteurs les assimilent à l’ordre des Blattodea. En effet les analyses phylogénétiques les plus récentes montrent que les termites forment un groupe monophylétique des Blattoptères et que leur groupe frère correspond aux blattes sub-sociales xylophages (qui se nourrit de bois) du genre Cryptocercus (famille des Cryptocercidae). Malgré la forte similitude avec les Hyménoptères eusociaux (les fourmis, certaines guêpes et abeilles), les termites ont un système de reproduction diplo-diploïde et les deux sexes coexistent au sein de chaque caste. Contrairement aux Hyménoptères qui sont Holométaboles, les termites sont des insectes Hétérométaboles ou encore Paurométaboles.
Les termites se composent de huit familles (Figure 1), avec notamment les Rhinotermitidae et les Termitidae qui constituent les lignées les plus apicales dans l’arbre phylogénétique, qui présentent le nombre d’espèces le plus important avec environ 300 et 2 000 espèces connues, respectivement.
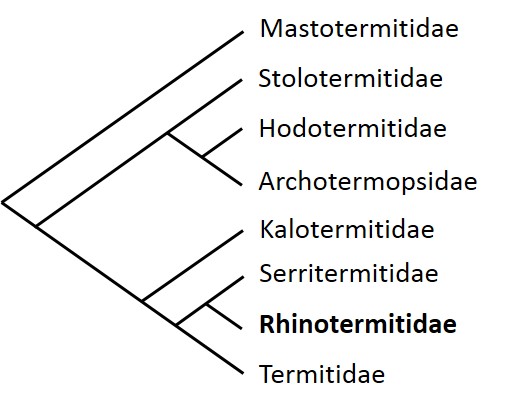
Ici, nous nous intéresserons principalement aux termites de la famille des Rhinotermitidae et plus particulièrement aux espèces du genre Reticulitermes.
Distribution géographiques & écologie des Reticulitermes
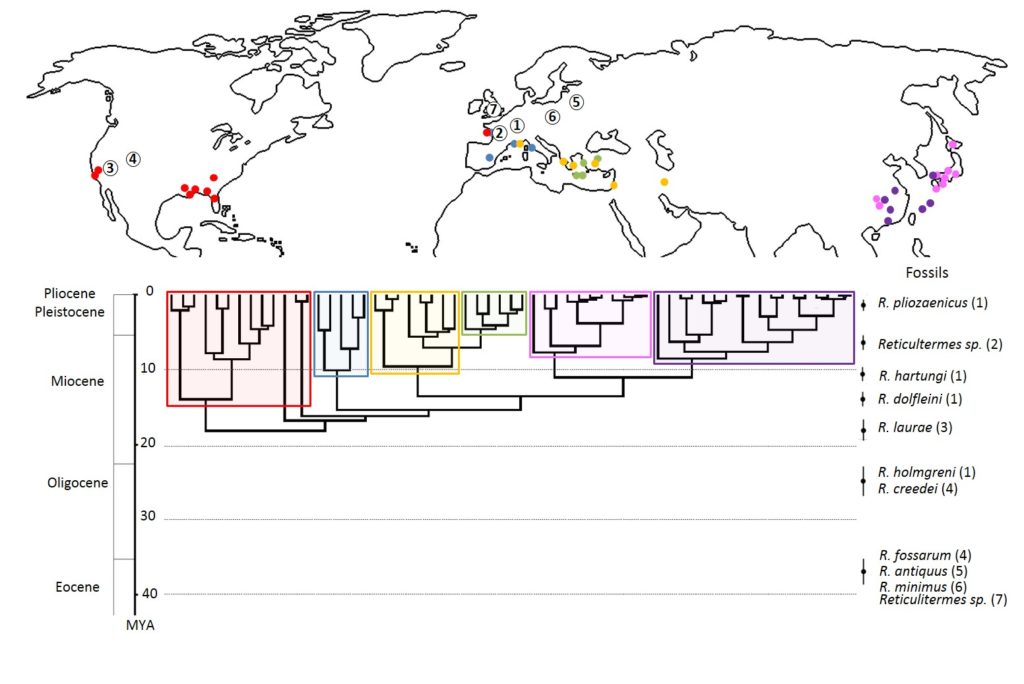
Les termites du genre Reticulitermes Holmgren (1913) appartiennent à la famille des Rhinotermitidae. On estime la diversification de ce genre à environ 38 millions d’années, où la plupart des espèces auraient pour origine le continent Nord-Américain. À ce jour, on compte environ 138 espèces, principalement réparties dans les régions tempérées du globe (Figure 2). Principalement situé dans l’hémisphère Nord. L’ensemble des espèces de ce genre sont dites « souterraines » ; elles s’alimentent de bois mort et construisent leurs nids dans le sol ou dans le bois. Dans la nature, les termites souterrains jouent ainsi un rôle écologique important dans la décomposition de la matière organique et l’aération des sols.
Ces termites lucifuges (qui fuit la lumière) construisent de véritables réseaux de galeries (d’où leur nom) souterraines et aériennes les protégeant de la dessiccation et leur permettant d’atteindre de nouvelles sources de nourritures. Durant la phase d’exploration, seuls quelques ouvriers explorent de nouveaux territoires dans toutes les directions et une fois la nourriture détectée grâces aux molécules volatiles qu’elle émet, les fourrageurs entreprennent des explorations plus longues et plus fréquentes en direction de la source de nourriture, et recrutent d’autres ouvriers pour l’exploitation de cette ressource. Cependant, en milieu urbain, les Reticulitermes peuvent être très destructeurs pour les infrastructures contenant du bois.
Impacts économiques
Le groupe des termites souterrains (Rhinotermitidae) est l’un des groupes de termites possédant le plus grand nombre d’espèces envahissantes, avec notamment les espèces des genres Coptotermes et Reticulitermes. À elles seules, les espèces de ces deux genres sont responsables de 80% des dommages attribués aux termites dans le monde. De par leur écologie (c.-à-d., une vie souterraine et un mode de nutrition xylophage), ces espèces infestent régulièrement les infrastructures humaines. Aux États-Unis entre autres, la lutte contre les termites et les dégâts qu’ils occasionnent coûtent des milliards de dollars chaque année (estimation de 32 milliards de dollars en 2010).
Description des espèces présentes en Europe
Sur les six espèces de Reticulitermes présentes en France, quatre sont natives d’Europe : on trouve sur le pourtour méditerranéen les deux sous-espèces R. lucifugus lucifugus et R. lucifugus corsicus et, provenant de la péninsule ibérique, les espèces R. grassei et R. banyulensis. Egalement deux espèces invasives sont présentes sur le territoire français, R. urbis et R. flavipes, respectivement originaires des Balkans et des États-Unis.
Ces espèces peuvent se retrouver en zones urbaines loin de leurs aires d’origine, comme par exemple R. banyulensis à Lyon ou R. lucifugus à Bordeaux, ou encore à Marseille où 5 des 6 espèces sont retrouvées. Récemment, des colonies de R. grassei ont été observées hors de leur aire de répartition dans la région de Tours (Projet TermiCentre 2012-2015) ; Reticulitermes balkanensis, espèce des Balkans proche de R. urbis, est la seule espèce européenne décrite à ce jour à n’avoir pas été trouvée sur le territoire français.
Organisation sociale
Les Reticulitermes, comme tous les termites, ont un développement incomplet, ce qui fait que chaque œuf pondu a un potentiel ontogénique (développement progressif d’un organisme) particulier, pouvant donner lieu à plusieurs issues développementales possibles. Les colonies de termites sont organisées en système de castes répartissant les individus sous différentes fonctions propres à la vie coloniale. Les différentes castes observées chez les termites du genre Reticulitermes ont toutes leur voie développementale particulière (lire cet article). Mais on peut dégager deux lignées principales : la lignée ouvrière, où les individus sont aptères (sans aile), et la lignée nymphale, où les individus sont brachyptères (avec développement de bourgeons alaires et d’ailes).
La lignée ouvrière comporte en principe des castes stériles. On y trouve la caste des ouvriers (Figures 4B) qui représente la majorité des individus de la colonie (environ 80%). Ils construisent le nid, recherchent de la nourriture, prennent soin des jeunes et nourrissent les autres individus de la colonie. On y trouve aussi la caste des soldats, issue de la différenciation d’ouvriers (Figure 4C). Ils assurent la défense de la colonie et représentent environ 2% de la colonie.
La lignée nymphale comporte principalement les nymphes (Figures 4D) dont les mues successives peuvent les différentier en imagos ailés, individus matures sexuellement (Figures 4F). Ces imagos, seuls adultes de la colonie, essaiment annuellement au printemps afin de rechercher un partenaire sexuel et fonder une nouvelle colonie. Ces reproducteurs sont appelés reproducteurs primaires : ce sont eux qui classiquement fondent et assurent la reproduction de la colonie. Il existe deux chemins développementaux pouvant aboutir à la différenciation de reproducteurs secondaires ou néoténiques (Figures 4E).
Les néoténiques sont de deux types : les néoténiques brachyptères (ou nymphoïdes), issus de la différentiation de nymphes, et les néoténiques aptères (ou ergatoïdes) issus de la différentiation d’ouvriers. Ces derniers n’ont jamais été observés dans la nature, mais sont régulièrement observés dans des colonies isolées en laboratoire. Ces reproducteurs secondaires viennent en remplacement ou en complément des reproducteurs primaires et ont une reproduction phylopatrique.

L’origine de la différenciation des néoténiques est encore très mal connue. On sait néanmoins que, chez l’espèce asiatique, R. speratus, la différenciation de néoténiques femelles est inhibée par des phéromones émises par les néoténiques femelles reproductrices et, à l’inverse, ces phéromones stimulent la différenciation de néoténiques mâles.
Très récemment, il a pu également être mis en évidence cette inhibition avec des néoténiques ergatoïdes femelles (issus de la différenciation d’ouvriers) chez R. flavipes. Par ailleurs, chez certaines espèces du genre, les femelles néoténiques se différencient à partir d’œufs non-fécondés de manière parthénogénétique.
Dispersion et fondation des nouvelles colonies
L’existence de ces deux types de reproducteurs (primaires et secondaires) est associée à deux modes de fondation coloniale possible chez les Reticulitermes ; (1) l’essaimage et (2) le bouturage.
L’essaimage est le mode de dispersion classique présent chez toutes les espèces de termites. Au printemps, les nymphes à longs fourreaux qui ont réalisé leur mue imaginale se mélanisent avant d’essaimer pour un vol nuptial. Au cours de l’essaimage, mâles et femelles partent à la recherche d’un partenaire pour fonder une nouvelle colonie. Le plus souvent, le vol sert principalement à éloigner les individus de la colonie dont ils sont issus, ce qui permet d’éviter les appariements consanguins. Le vol nuptial n’excède généralement pas une centaine de mètres et correspond donc à la distance de dispersion par essaimage.
Une fois au sol, les mâles recherchent une femelle activement. Cette recherche est principalement orientée grâce aux phéromones volatiles émises par les femelles qui permettent aux mâles de pouvoir les retrouver. Cependant, l’appariement entre les individus ne semble pas tenir compte de l’apparentement entre les individus et se produit aléatoirement. Ensuite, les partenaires restent en couple monogame, copulent et partent ensemble à la recherche d’un site favorable pour y fonder une nouvelle colonie
Le bouturage est a priori moins répandu chez les termites, et bien qu’il n’ait jamais été démontré formellement dans la nature, il a régulièrement été suspecté dans de nombreuses études des populations de Reticulitermes. Ce mode de dispersion est assuré via la différenciation de reproducteurs secondaires. En effet, lorsque les colonies disposent d’abondantes ressources alimentaires à proximité des nids, la présence de néoténiques permet l’accroissement de l’exploitation de cette ressource. Comparer au nombre de descendants produits par une seule reine, la présence de néoténiques femelles aura tendance à accroître la productivité de la colonie. Cependant, les ouvriers et nymphes ont tendance à rechercher de nouvelles sources de nourriture aux alentours de leur nid constitué de tout un réseau de galeries, et il arrive que certains de ces individus se différencient en reproducteurs néoténiques, créant ainsi un réseau de nids satellites.
Par ce phénomène, les colonies de termites peuvent s’étendre de proches en proches, et ce parfois sur plusieurs centaines de mètres, voire dans certains cas, jusqu’au kilomètre. Dans de telles situations, il pourrait arriver qu’un nid satellite se retrouve isolé des autres nids et forme une nouvelle entité coloniale indépendante de la colonie parentale. Ce mode de fondation peut avoir lieu sur de courtes distances comme on vient de le voir, mais aussi, avec l’aide de l’homme, sur de longues distances. Plusieurs études ont d’ailleurs révélé que quelques individus seulement (une cinquantaine d’ouvriers ou de nymphes) suffisent à la fondation d’une nouvelle colonie via la différenciation en néoténiques. De façon accidentelle, l’Homme peut ainsi contribuer à la dispersion longue-distance de ces espèces. Le bouturage a ainsi été proposé comme étant un mode de dispersion probable en milieu urbain.

Système de dispersion
L’organisation sociale des colonies de termites peut prendre différentes formes en fonction du nombre, du type (primaire ou secondaire), du sex-ratio et des liens de parentés parmi les reproducteurs.
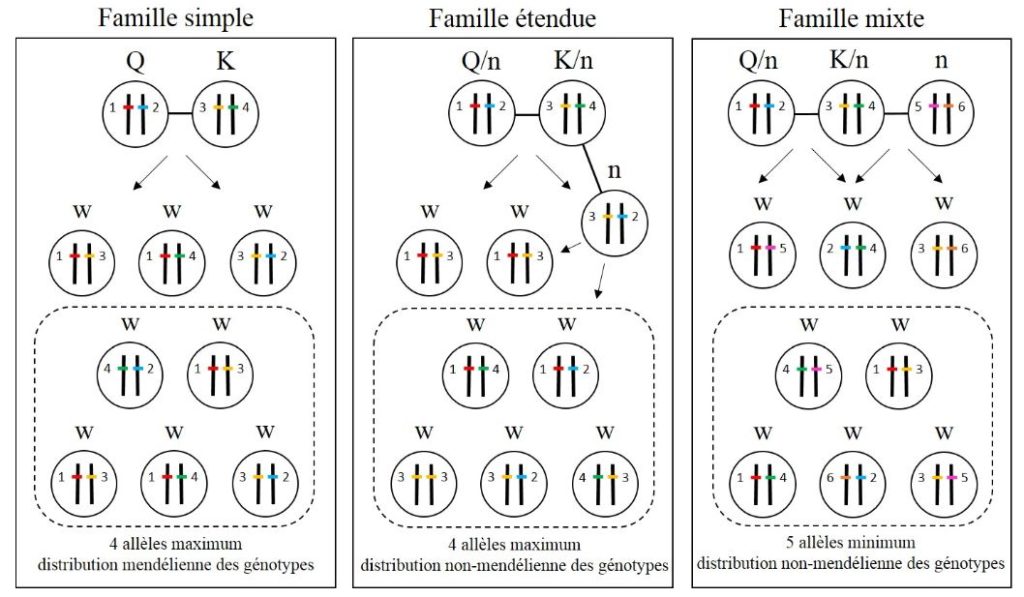
De par leur mode de vie cryptique, les reproducteurs des termites souterrains, comme les Reticulitermes, ne sont généralement pas accessibles. L’utilisation de l’outil moléculaire permet, par la collecte d’une dizaine/vingtaine d’ouvriers à un ou plusieurs sites de fourragement, d’identifier les limites des colonies et la structure reproductrice des colonies. Grâce aux marqueurs génétiques comme les microsatellites, il est possible d’identifier le nombre de reproducteurs, ainsi que les liens de parentés des individus au sein des colonies en déterminant la distribution des génotypes et des fréquences alléliques dans la descendance des reproducteurs (Figure 5). Ainsi, trois grands types de familles ont été définis chez les termites souterrains.
La famille simple est la structure familiale la plus communément observée chez les termites souterrains. Elle est composée par le couple primaire monogame, le roi et la reine (généralement non apparentés), et leurs descendants stériles. Ainsi, l’analyse génétique d’une famille simple ne révèle jamais plus de 4 allèles à chacun des loci étudiés. De plus, les fréquences génotypiques attendues dans la descendance sont a priori conformes aux proportions mendéliennes (Figure 5)
La famille étendue est une structure familiale qui, en remplacement ou en supplément des deux reproducteurs primaires, est le fruit de la reproduction consanguine de reproducteurs secondaires néoténiques. Comme dans les familles simples, on observe au maximum 4 allèles à tous les loci considérés, ce qui représente le maximum d’allèles que peuvent porter les deux reproducteurs primaires à l’origine de la colonie. Par contre, la distribution des génotypes dans la descendance d’une famille étendue n’est pas conforme à une distribution mendélienne (Figure 5). En effet, avec la reproduction consanguine des néoténiques, il est attendu que le degré de parenté entre descendants soit plus important que dans une famille simple, et qu’il augmentera avec le nombre de néoténiques fertiles au sein de la colonie, ceci ayant pour conséquence une distribution non-mendélienne des génotypes dans la descendance. Ainsi, dans les colonies de ce type familial. Dans la majorité des cas, cependant, nous n’avons pas accès aux reproducteurs, et on ne peut dès lors pas exclure que seuls des néoténiques restent au sein de la colonie et se reproduisent entre eux, et ce, sur plusieurs générations consanguines qui se chevauchent.
La famille mixte est une structure familiale issue de la reproduction de plus de deux reproducteurs non apparentés. Chez les termites, cette structure familiale peut être obtenue de plusieurs manières :
-
- Par l’adoption de nouveaux reproducteurs dans une colonie mature
-
- Par la pléométrose, qui est la fondation d’une colonie par plus de deux reproducteurs non-apparentés (polygamie), le plus souvent un mâle et deux femelles
- Par la fusion de deux colonies. Chez les Reticulitermes, seule la fusion coloniale a été observée pour former une famille mixte. Au sein de ces colonies, les reproducteurs primaires et/ou secondaires, provenant des deux colonies non-apparentées, peuvent se reproduire ensemble. On peut facilement identifier ce type familial avec l’utilisation de marqueurs génétiques lorsque, à au moins un locus sont relevés un minimum de 5 allèles différents. Cependant, la détection de ces familles mixtes reste dépendante du taux de polymorphisme des loci utilisés dans la population étudiée

Bien qu’il soit difficile de connaitre, par observation directe, la nature des reproducteurs présents dans les colonies, il est possible d’estimer le nombre de reproducteurs au sein des colonies grâce à l’utilisation des F-statistiques hiérarchiques. En comparant les F-statistiques calculées avec les colonies étudiées et les F-statistiques théoriques obtenues par des systèmes de reproduction simulés, il est possible d’estimer le nombre de reproducteurs présents au sein des colonies étudiées. La présence de néoténiques dans les colonies va augmenter les niveaux de consanguinité individuelle (FIC), et l’appariement de reproducteurs non apparentés (exemple des familles mixtes) aura tendance à diminuer l’apparentement (r) des individus au sein des colonies.
Il est connu dans la littérature que certaines espèces du genre Reticulitermes sont également capables de produire des descendants par parthénogénèse thélytoque (reproduction à partir de gamètes (haploïdes) femelles non fécondées donnant uniquement des femelles (diploïdes)). Le mécanisme mis en jeux au cours de cette reproduction asexuée chez les Reticulitermes est l’automixie avec une fusion terminale (fusion de 2 noyaux haploïdes après la deuxième division de méiose).
Chez ces espèces, les reines utilisent exclusivement la parthénogénèse pour produire leurs filles néoténiques, bien que ces dernières aillent se reproduire de manière sexuée, en s’accouplant avec le mâle primaire. On nomme ce système de reproduction Asexual Queen Succession (AQS). L’ensemble des descendants produits par parthénogénèse chez ces espèces sont des femelles néoténiques.
Les individus homozygotes à tous les loci étudiés (microsatellites) sont considérés comme étant des produits de la parthénogénèse. Donc en analysant le taux d’homozygotie chez les individus néoténiques femelles, il est possible de pouvoir identifier les espèces capables de parthénogénèse thélytoque. À ce jour, la présence d’AQS a été mise en évidence chez trois espèces de Reticulitermes : R. speratus, R. virginicus et R. lucifugus.
Reticulitermes flavipes : histoire d’une invasion
Distribution géographique de R. flavipes
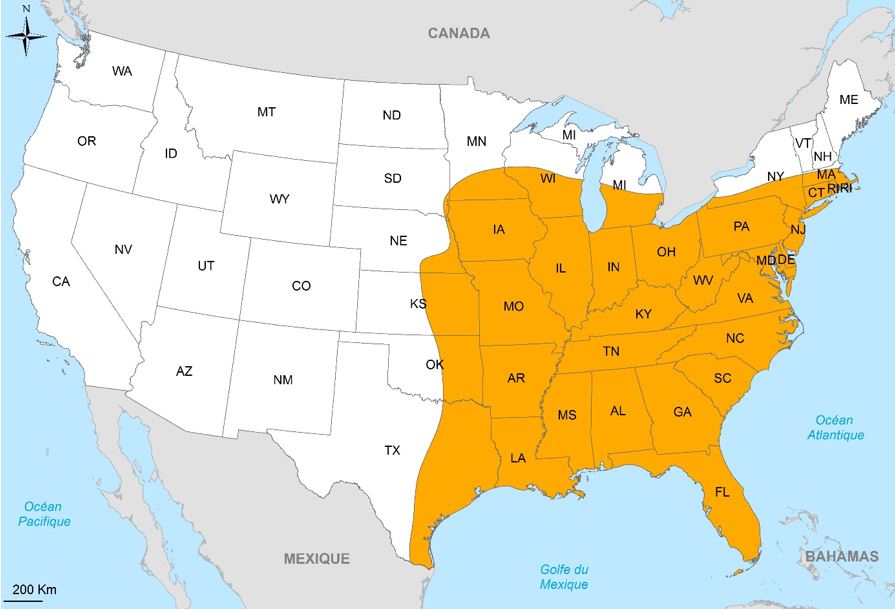
Le termite à patte jaune, Reticulitermes flavipes Kollar, est une espèce originaire des États-Unis d’Amérique. Son aire de répartition couvre une grande partie de l’est et du sud-est des États-Unis (Figure 6). Bien que d’autres espèces de termites souterrains natifs vivent en sympatrie avec cette espèce (R. virginicus, R. hageni, R. hesperus, R. malletei, R. tibialis et R. nelsonae), R. flavipes est l’une des espèces causant le plus de dommages aux États-Unis.
Cette espèce est très invasive et a été introduite dans plusieurs régions du monde. Aux États-Unis, elle a été introduite dans d’autres états que ceux du grand Est, tels que la Californie ou le Nebraska, ainsi que dans d’autres pays d’Amérique du Nord (Canada et Bahamas) et du Sud (Chili et Uruguay).
En Europe, elle est largement répandue en France (Figure 8) et a été ponctuellement retrouvée en Allemagne (Hambourg), en Italie (Varèse) et en Autriche (Vienne), pays où elle a été décrite pour la première fois par Kollar. En France, bien que cette espèce fût longtemps considérée comme européenne et appelée R. santonensis, ou termite de Saintonge, plusieurs études contemporaines s’accordaient sur le fait que cette « espèce » était en réalité une population introduite de R. flavipes depuis les États-Unis.
Une analyse de plus de 200 échantillons collectés aux États-Unis et en France a révélé l’appartenance génétique de la population française à l’espèce américaine R. flavipes. Cette étude a également révélé que l’ensemble des infestations françaises échantillonnées sont originaires d’une population source américaine de l’état de Louisiane et plus précisément de la région de la ville de la Nouvelle-Orléans.
Variabilité des systèmes de reproduction
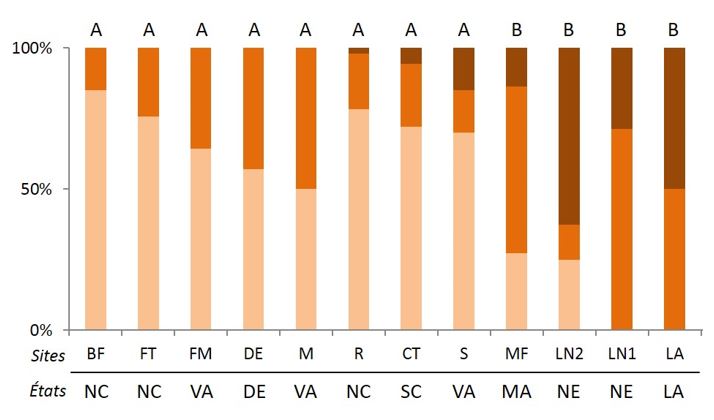
Ce termite a fait l’objet, dans son aire native, de nombreuses études, notamment sur les systèmes de reproduction qui ont été particulièrement bien caractérisés. Ces études ont montré qu’il y a une grande variabilité dans les proportions des différents types de familles observés chez cette espèce. Même si, dans son aire native, les colonies observées sont majoritairement des familles simples, la proportion de familles étendues et mixtes, ainsi que le nombre de néoténiques au sein des colonies sont variables (Figure 7).
Une étude récente de Vargo et al. (2013) a pu mettre en avant que les variations des systèmes de reproduction des colonies aux États-Unis étaient corrélées à des variations latitudinales. En effet, l’apparition de familles étendues et mixtes, et surtout de fort taux de consanguinité, ainsi qu’un plus grand nombre de reproducteurs, favoriseraient la persistance des colonies dans des latitudes plus élevées au Nord, où les températures sont plus fraîches et l’humidité du sol plus importante.
De plus, les variations saisonnières favoriseraient les colonies de grandes tailles durant la période froide et seraient limitantes pour les colonies de type famille simple, notamment lors des premières étapes de la fondation coloniale par les ailés. La grande taille de ces colonies et leur taux de consanguinité élevé sont assurés par la production d’un très grand nombre d’œufs issus de multiples néoténiques femelles. Cependant, la découverte dans la ville de la Nouvelle Orléans en Louisiane (Figure 7) d’un très grand nombre de colonies étendues et mixtes possédant de multiples néoténiques suggère que d’autres facteurs, tel que le milieu urbain, peuvent façonner la structure reproductrice chez cette espèce.
La suite lors d’un prochain article…
Interview de l’auteur
 Bien que les termites causent de nombreux dégât dans le patrimoine parisien, peu de choses sont connues sur leurs modes d’expansion dans le milieu urbain. Il était donc nécessaire de mieux comprendre leur écologie dans le but prédire leur expansion future et donc anticiper les moyens de préventions et de luttes à mettre en place. Le fait que les termites soient des insectes sociaux avec une structure de fonctionnement complexe a vraiment suscité de l’intérêt pour un généticien des populations comme moi.
Mon travail a consisté à collecter un maximum d’individus sur le territoire parisien afin d’identifier les différentes populations et colonies présentes dans la région afin de retracer les liens de parenté existant entres elles. L’objectif était d’identifier les voies (à travers le paysage parisien) utilisées par les termites pour infester de nouveaux bâtiments et de nouveaux quartiers. Mes recherches ont permis d’appliquer et d’adapter une nouvelle discipline : la génétique du paysage, qui associe génétique des populations et écologie du paysage. L’utilisation de ces approches a permis d’identifier la voie ferrée comme un élément clé du paysage dans la dissémination des termites, tant à Paris qu’à l’échelle de la France.
Je pense que les gens ne sont pas assez informés sur cette problématique urbaine existante depuis 1945 à Paris. Récemment (1999) une loi permet de protéger les acquéreurs contre cette problématique mais cela n’est pas suffisant pour limiter leur propagation, quasiment uniquement imputée aux activités humaines. L’espèce sur laquelle je travaillais, Reticulitermes flavipes, est originaire des USA où l’espèce occasionne également de nombreux dégâts. R. flavipes se rencontre dans son aire d’origine aussi bien en forêt qu’en ville, en France, l’espèce est, quant à elle, principalement présente en ville. Cependant, des populations se développent dans les forêts côtières entre la Vendée et la Charente. Le rôle écologique des termites en forêt est majeur, ils dégradent la matière organique (bois et végétaux) et participent ainsi au renouvellement, la fertilisation et à l’aération des sols.
Non pas vraiment, il s’agit de bien informer le public pour que chacun prenne conscience et soit acteur dans cette problématique. Les efforts de protection doivent être mutualisés : si, seul un des voisins réalise un traitement anti-termites, il existe une grande probabilité pour que les autres habitations soient infestées ou ré-infestées dans les années qui suivent.
Pas une anecdote, mais un moment qui m’a beaucoup plu. Avec mes collègues de la Mairie de Paris, nous avions été informés de la présence de termites près de l’observatoire de Paris. L’observatoire possédant un grand terrain constitué de bois morts et de branches au sol, propice à la présence de termites, nous avons demandé à le visiter. En discutant, le responsable nous a indiqué l’existence, sous les jardins du parc, de nombreux souterrains non accessibles au public. Nos termites étant souterrains, il nous a donc été proposé de visiter exceptionnellement les lieux. Bien qu’aucun termite n’ait pu être rencontré, nous avons découvert tout un réseau de salles d’expérimentation où étaient entreposés des horloges à pile et un thermomètre de Lavoisier datant de la Révolution ! L’insurgé pensait du Comte de Cassini cachait des nobles dans ses souterrains. |
Sources
-
- Bankhead-Dronnet S. ; Perdereau E. ; Kutnik M. ; et al (2015) Spatial structuring of the population genetics of a European subterranean termite species. Ecol Evol 5:3090–3102
-
- Brune A. (2014) Symbiotic digestion of lignocellulose in termite guts. Nat Rev Microbiol 12:168–180
-
- Bulmer M.S. ; Adams E.S. ; Traniello J.F.A. (2001) Variation in colony structure in the subterranean termite Reticulitermes flavipes. Behav Ecol Sociobiol 49:236–243
-
- Dedeine F. ; Dupont S. ; Guyot S. ; et al (2016) Historical biogeography of Reticulitermes termites (Isoptera: Rhinotermitidae) inferred from analyses of mitochondrial and nuclear loci. Mol Phylogenet Evol 94:778–790
-
- DeHeer C.J. ; Vargo E.L. (2004) Colony genetic organization and colony fusion in the termite Reticulitermes flavipes as revealed by foraging patterns over time and space. Mol Ecol 13:431–441
-
- Misof B. ; Liu S, Meusemann K. ; et al (2014) Phylogenomics resolves the timing and pattern of insect evolution. Science 346:763–767
-
- Perdereau E. ; Bagnères A.G. ; Vargo E.L. ; Baudouin G. ; et al (2015) Relationship between invasion success and colony breeding structure in a subterranean termite. Mol Ecol 24:2125–2142 (lien)
-
- Vargo E.L. (2000) Polymorphism at trinucleotide microsatellite loci in the subterranean termite Reticulitermes flavipes. Molecular-Ecology 9:817–829
- Vargo E.L., Leniaud L. ; Swoboda L.E. ; et al (2013) Clinal variation in colony breeding structure and level of inbreeding in the subterranean termites Reticulitermes flavipes and R. grassei. Mol Ecol 22:1447–1462
Articles de l’auteur
- Baudouin G. ; Dedeine F. ; Bech N. ; Bankhead-Dronnet S. ; Dupont S. (2017) : An American termite in Paris : temporal colony dynamics. Genetica, vol145(6):491-502 (lien)
- Baudouin G. ; Bech N. ; Bagnères A.G. ; Dedeine F. (2018) : Spatial and genetic distribution of a north American termite, Reticulitermes flavipes, across the landscape of Paris. Urban Ecosystems, V21(4):751-764 (lien)





Bonjour merci pour cet article.
Svp je travaille sur l’évaluation de la fréquence de patrouille en fonction du volume du nid chez certains termitinae pouvez vous me donner quelques éléments ?
Merci bien