Ecologie nutritionnelle des herbivores
Par Marion Le Gall
Depuis mon enfance, j’ai toujours été fascinée par les animaux de petite taille. Le jardin public adjacent à notre maison m’offrait une alternative naturelle à la collection grandissante de Polly Pocket qu’abritait le tiroir sous mon lit. Je passais des heures à attraper têtards, tritons, invertébrés aquatiques, orthoptères, et coléoptères. De cette époque dans ma Bretagne natale, mes premiers livres d’entomologie me suivent toujours et se trouvent à présent chez moi dans le désert du Sonora en Arizona, au côté d’autres livres d’entomologie tels que ma précieuse copie du « Voyage chez les fourmis », un cadeau de Bert Hölldobler.

Parmi ces livres d’enfance, un détail amusant m’est revenu il y a peu. Le livre en question1, probablement un cadeau de mon père, présente un magnifique scarabée tenant un registre sur lequel le nom du propriétaire de l’ouvrage peut être inscrit. Sur cette page on peut distinguer mon nom inscrit dans l’écriture paternelle mais aussi, en dehors du format prescrit, le prénom de ma sœur. Cette insertion, probablement le fait d’une rivalité enfantine (ma sœur ne possède pas mon intérêt pour les insectes) a dû provoquer chez moi un sentiment vindicatif puisque l’addition épistolaire est gribouillée et remplacée par ma propre signature. Ou son équivalent. Mon amour des insectes précède ma commande de l’écriture, j’ai donc inscrit une série de lettres « Emlmion » qui dans mon esprit suffisaient probablement à rétablir mon autorité sur le précieux ouvrage (Figure 1).
Seize ans plus tard, sachant finalement écrire, je quittais la Bretagne pour Tours afin de poursuivre ma dernière année de licence et mon master au sein de l’Institut de Recherche sur la Biologie de l’Insecte à l’Université François-Rabelais (IRBI). Durant mon master, j’ai travaillé sur deux projets de recherche : l’un sur le mimétisme chez une espèce d’araignée crabe (Misumena vatia) dans l’équipe de Jérôme Casas et l’autre sur la fonction antioxydante des pigments présents dans les yeux de punaises hématophages (Rhodnius prolixus) dans l’équipe de Claudio Lazzari 2. Cette introduction au travail de recherche à l’IRBI a été déterminante dans mon choix de poursuivre une carrière dans la recherche entomologique. C’est pourquoi, six mois après l’obtention de mon master, je me suis retrouvée dans un avion direction le Texas pour effectuer ma thèse dans le laboratoire de Spencer Behmer à Texas A&M University.
Ayant précédemment très peu voyagé, je fus surprise des dissimilarités entre le système universitaire français et le système américain. Aux Etats-Unis, même les universités publiques sont payantes.
Par exemple, en 2019/2020 les frais de scolarité à Texas A&M University étaient de $13,012 par an pour une licence et $8,473 pour un master/thésard (https://financialaid.tamu.edu/). Ces coûts peuvent être pris en charge par l’université pour les étudiants qui obtiennent une position d’enseignant de travaux dirigés à raison de 20h par semaine. Cela a été mon cas. Pendant quatre semestres, j’ai enseigné une introduction à l’entomologie pour les étudiants en licence de deuxième année, et pendant deux semestres une classe sur la physiologie de l’insecte pour des étudiants en troisième ou quatrième année de licence (lien).
Chuck Norris mis à part, j’avais choisi le laboratoire de recherche au Texas parce que mon intérêt pour l’entomologie était devenu plus spécifique durant mon master. J’étais (et suis toujours) fascinée par la façon dont les insectes interagissent avec leur environnement. Sous la houlette de mon maitre de thèse, je me suis spécialisée dans l’écologie nutritionnelle des herbivores. Quelques années avant le début de ma thèse, une étude était sortie qui avait démontré que le contenu en protéines et en glucides de la nourriture pouvait mitiger les effets des toxines présentes dans les plantes pour une espèce de criquet migrateur (Locusta migratoria)3.
Cette étude utilisait une approche élégante, développée en Australie dans les années 90 par David Raubenheimer et Steve Simpson, qui permet de séparer les effets caloriques des nutriments de leur ratio en représentant les nutriments dans un espace géométrique (Geometric Framework for Nutrition, 4–7).
Cette approche de géométrie nutritionnelle avait permis de démontrer, à l’origine chez les criquets, puis par la suite pour une vaste série d’organismes, que les animaux peuvent réguler leurs apports en nutriments. Par exemple, en donnant à un criquet le choix entre deux nourritures, une riche en protéines et l’autre en glucides, on peut s’apercevoir que la consommation entre les deux nourritures n’est pas aléatoire et que les criquets choisissent un ratio spécifique (Figure 2).
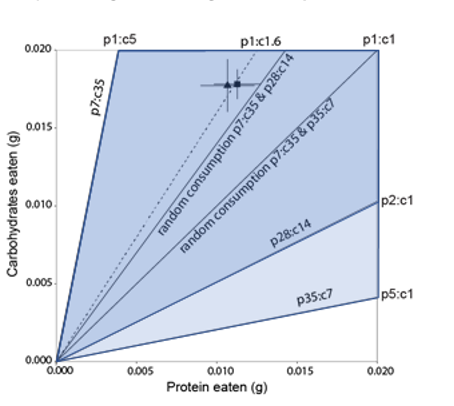
Pour chacun des deux traitements, si la consommation entre les deux nourritures est aléatoire, les points devraient se trouver au milieu sur les lignes de consommation aléatoire (random consumption). On peut voir que ça n’est pas le cas et que pour les deux traitements les criquets choisissent le même ratio de protéines et de glucides (proteine 1 : glucides 1.6) (Source : Adapté de Le Gall et al., Functional Ecology, 2019)
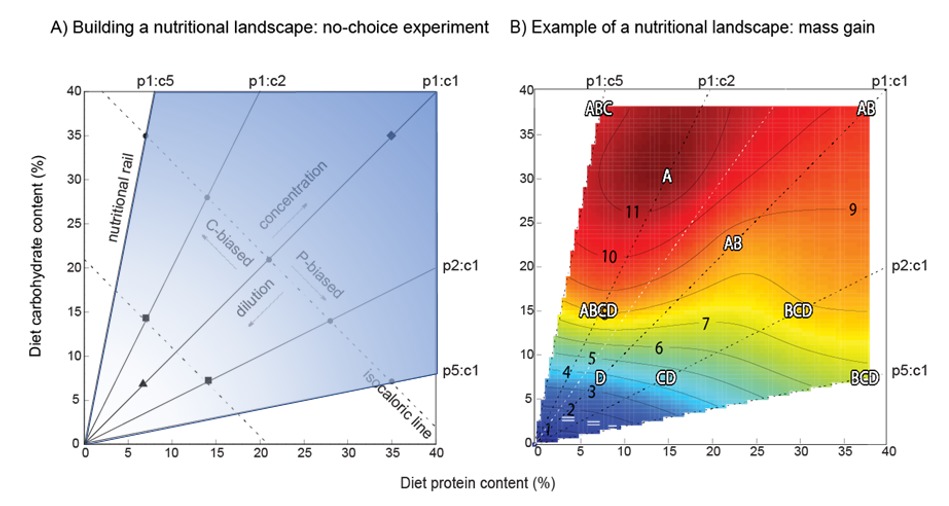
En l’absence de choix, quand les criquets mangent une nourriture présentant un ratio optimal de protéines et de glucides ils se développent plus rapidement, deviennent plus gros, et sont en générale plus performants8,9 (Figure 3).

Durant ma thèse, j’ai donc utilisé la méthode de la géométrie nutritionnelle pour évaluer les effets des toxines sur le choix et la performance d’une espèce de criquet largement répandu aux Etats-Unis, Melanoplus differentialis. C’est une espèce d’une belle couleur jaune facilement identifiable grâce aux marques qui forment un motif de chevron sur les pattes arrière (Figure 4).
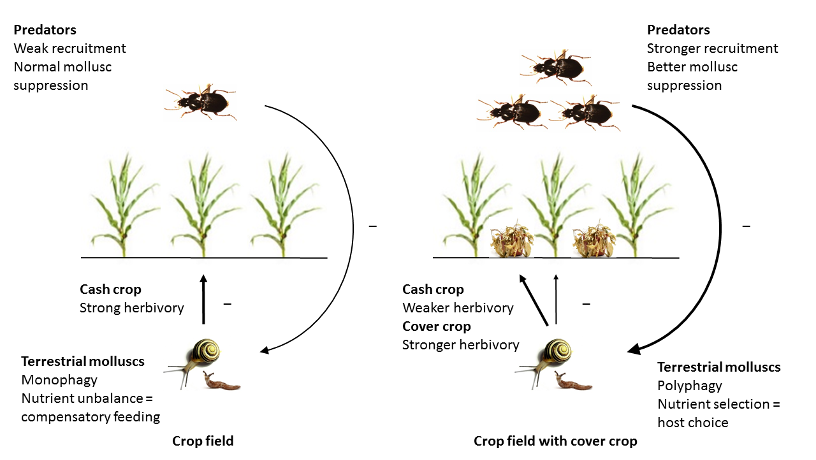
Une fois ma thèse finie, je suis restée aux Etats-Unis. Direction cette fois le Nord-Est pour rejoindre le département d’entomologie à Penn State University en Pennsylvanie. Un petit voyage de 2400 kilomètres. En travaillant avec mon chef, John Tooker, je suis passée des criquets aux… limaces. L’objectif étant d’utiliser l’approche de la géométrie nutritionnelle pour 1) comprendre les préférences de ces animaux problématiques pour les cultures et 2) développer un programme de management basé sur leur biologie10.
Ce postdoc fut également pour moi une opportunité de travailler dans l’entomologie agriculturale et d’adapter mes connaissances basiques acquises en écologie nutritionnelle à des applications pratiques (Figure 5).
En tant qu’entomologiste, malgré ma fascination et mon respect pour ces petits mollusques, les limaces me donnaient quelque peu une crise identitaire. Heureusement pour moi, pendant ma première année de thèse mon chemin avait croisé celui d’une autre thésarde, Arianne Cease, qui revenait à l’époque de Chine ou elle avait collecté des données sur la nutrition du criquet asiatique (Oedaleus asiaticus) qui menèrent a une publication pour le prestigieux journal « Science »11. Elle avait depuis monté un laboratoire de recherche sur les criquets migrateurs en Arizona : Global Locust Initiative Lab. Alors que la fin de mon postdoc à Penn State University approchait, je l’ai approchée lors d’un séminaire pour lui demander si elle cherchait un postdoc pour travailler sur les criquets migrateurs. Un déménagement de plus de 3500 kilomètres cette fois (trois fois Brest-Moscou) et me voilà donc en Arizona, dans le magnifique désert du Sonora.
Non contente d’avoir monté un laboratoire de recherche classique, Arianne avait aussi créer une initiative pour rassembler tous les partenaires travaillant sur la lutte contre les criquets migrateurs aux quatre coins du monde, depuis les agriculteurs, agences gouvernementales, jusqu’aux scientifiques (Global Locust Initiative, https://sustainability-innovation.asu.edu/global-locust-initiative/). C’est grâce à ces connections avec des partenaires locaux, que je me suis envolée pour le Sénégal pour la première fois en 2017, puis deux fois en 2018, et trois fois en 2019. Pour faire bonne mesure, j’ai également effectué une saison de terrain en Australie en 2017.
Un des résultats surprenant de mon travail au Sénégal sur l’écologie nutritionnelle du criquet sénégalais12,13 est leur préférence pour les glucides plutôt que les protéines. Des résultats similaires ont été établis pour d’autres espèces de criquets migrateurs comme le criquet asiatique (O. asiaticus)11,14, le criquet australien (Chortoicetes terminifera)15, le criquet sud-américain (Schistocerca cancellata)16, et d’autres espèces migratrices9.
Cela est surprenant puisque la plupart des herbivores ont tendance à préférer, et à démontrer de meilleures performances, quand la teneur en protéines de leur nourriture est haute. Cette règle tient du fait que les plantes sont généralement limitées en apport azotés et contiennent moins de protéines que les tissus animaux17.
Cette préférence des criquets migrateurs pour les plantes plus pauvres en protéines offre des possibilité de lutte écologique en utilisant des techniques agriculturales qui visent à augmenter le contenu en azote du sol, ou à limiter l’érosion due à de trop haute densité de bétail dans les zones de pâturages14,18,19. Sachant que les pullulations de criquets coûtent des millions d’euros en pesticides et causent régulièrement des famines, pouvoir maintenir les criquets a de faibles densités en leur fournissant des plantes de qualités suboptimales est un aspect fondamental de notre recherche. L’autre aspect positif d’une approche préemptive est qu’elle permet de conserver les criquets dans leurs écosystèmes respectifs puisqu’ils sont des acteurs importants de la chaine alimentaire et participent au cycle des nutriments.
Mais pourquoi les criquets préfèrent-t-ils les glucides ?

Une des hypothèses regarde leurs capacités migratrices. Les longs voyages des criquets dépassent parfois plusieurs centaines de kilomètres par jour, ce qui représente un énorme coût calorifique. Les criquets utilisent généralement leurs réserves lipidiques pour assurer ces longs vols. Et quel meilleur moyen pour s’engrosser que de manger des glucides en excès! Donc, tout comme un coureur consomme une platée de nouille avant un marathon, nos criquets préfèrent les pâtes au poulet (article en Anglais : lien)
Une autre partie importante de mon travail au Sénégal a été la mise en place d’un système d’alerte précoce pour le criquet sénégalais. Grâce à un financement obtenu par Arianne auprès de l’Agence des États-Unis pour le développement international (USAID) (Article en Anglais : lien), et avec le support de la main d’œuvre locale, nous avons développés des pièges lumineux pour les criquets migrateurs. Nous avons placé ces pièges dans cinq villages localisés dans la zone la plus important de production de millet au Sénégal (communément appelée Bassin arachidier) et offert des formations à l’identification des criquets pour des comités de femmes (Figure 6). Les femmes de chaque village reportent directement leurs résultats à la Division de la Protection des Végétaux du Sénégal (lien) qui est l’agence gouvernementale avec laquelle nous travaillons pour ce projet (mais aussi le travail d’écologie nutritionnelle) et qui monitore et traite les pullulations.
Je voudrais remercier tous mes collègues et partenaires qui ont facilité cette recherche : les membres de l’IRBI, les membres de Texas A&M University, les membres de Penn State University, les membres d’Arizona State University. Je tiens à reconnaitre que le campus de Texas A&M University se trouve sur des terres prises au Tonkawa, Tawakoni, Hueco, Sana, Wichita, et Coahuiltecan ; le campus de Penn State University sur des terres prises aux Erie, Haudenosaunee, Lenni Lenape, Shawnee, Susquehannock, et Wahzhazhe Nations ; le campus d’Arizona State University se trouve sur des terres prises aux Akimel O’odham et Pee Posh. Je tiens également à remercier la Direction de la Protection des Végétaux au Sénégal et le Department of Primary Industries et l’Australian Plague Locust Commission pour leur support en Australie. Finalement, je voudrais remercier pour leur accueil les communautés de Trangie, Coonamble, Ivanhoe en Australie et les communautés de Nganda, Gnibi, Kaffrine, Mbar, Gossas au Sénégal.
Bibliographie
- Du Chatenet, G. Le livre des insectes. (Gallimard, 1989).
- Insausti, T. C., Le Gall, M. & Lazzari, C. R. (2013) : Oxidative stress, photodamage and the role of screening pigments in insect eyes. Journal of Experimental Biology 216, 3200–3207 (lien)
- Simpson, S. J. & Raubenheimer, D. (2001) : The geometric analysis of nutrient-allelochemical interactions: a case study using locusts. Ecology 82, 422–439 (lien)
- Raubenheimer, D. & Simpson, S. J. (1993) : The geometry of compensatory feeding in the locust. Animal Behaviour 45, 953–964 (lien)
- Raubenheimer, D. & Simpson, S. J. (1999) : Integrating nutrition: a geometrical approach. in Proceedings of the 10th International Symposium on Insect-Plant Relationships 67–82 (lien)
- Simpson, S. J. & Raubenheimer, D. (1993) : A multi-level analysis of feeding behaviour: the geometry of nutritional decisions. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 342, 381–402 (lien)
- Simpson, S. J. & Raubenheimer, D. (1995) : The geometric analysis of feeding and nutrition: a user’s guide. Journal of Insect Physiology 41, 545–553 (lien)
- Le Gall, M. & Behmer, S. T. (2014) : Effects of Protein and Carbohydrate on an Insect Herbivore: The Vista from a Fitness Landscape. Integrative and Comparative Biology 54, 942–954 (lien)
- Behmer, S. T. (2009) : Insect herbivore nutrient regulation. Annual review of entomology 54, 165–187 (lien)
- Le Gall, M. & Tooker, J. F. (2017) : Developing ecologically based pest management programs for terrestrial molluscs in field and forage crops. Journal of Pest Science 1–14 (lien)
- Cease, A. J. et al. (2012) : Heavy livestock grazing promotes locust outbreaks by lowering plant nitrogen content. Science 335, 467–469 (lien)
- Le Gall, M. et al. (2020) : Linking land use and the nutritional ecology of herbivores: A case study with the Senegalese locust. Functional Ecology 34, 167–181 (lien)
- Le Gall, M., Word, M. L., Thompson, N., Beye, A. & Cease, A. J. (2020) : Nitrogen fertilizer decreases survival and reproduction of female locusts by increasing plant protein to carbohydrate ratio. Journal of Animal Ecology 89, 2214–2221 (lien)
- Cease, A. J. et al. (2015) : Living with locusts: connecting soil nitrogen, locust outbreaks, livelihoods, and livestock markets. Bioscience 65, 551–558 (lien)
- Lawton, D., Le Gall, M., Waters, C. & Cease, A. J. (2021) : Mismatched diets: defining the nutritional landscape of grasshopper communities in a variable environment. Ecosphere 12, e03409 (lien)
- Talal, S. et al. (2020) : Plant carbohydrate content limits performance and lipid accumulation of an outbreaking herbivore. Proceedings of the Royal Society B 287, 20202500 (lien)
- White, T. C. The inadequate environment: nitrogen and the abundance of animals. (Springer Science & Business Media, 2012).
- Word, M. L. et al. (2019) : Soil-targeted interventions could alleviate locust and grasshopper pest pressure in West Africa. Science of The Total Environment (lien)
- Le Gall, M., Overson, R. & Cease, A. J. (2019) : A global review on locusts (Orthoptera: Acrididae) and their interactions with livestock grazing practices. Frontiers in Ecology and Evolution 7, 263 (lien)
Ouvrages sur la thématique
- Histoire d’un siècle de lutter anti-acridienne en Afrique : Contributions de la France (Jean Roy – Editions L’Harmattan – 296 pages – 1 janvier 2001)
- Le devenir des pesticides utilisés dans la lutte contre les acridiens: Le devenir des pesticides communément utilisés en lutte antiacridienne dans les aliments et dans les pâturages au Sahel (Hadya Mamoudou ; Bouaichi Abdelghani & Ahmed Mazih – Editions Universitaire européennes – 92 pages – 27 mai 2020)
- Sauterelles, grillons et criquets d’Europe occidentale (Hans Bellmann ; Florin Rutschmann ; Christian Roesti & Axel Hochkirch – Edition Delachaux – 320 pages – 25 juin 2020)





