Origine des ailes chez les insectes : le mystère en partie dévoilé !
Par Benoît GILLES
L’histoire évolutive de l’origine des ailes chez les insectes fascine les chercheurs depuis plus de 130 ans. Deux théories concurrentes expliquent leur apparition (lien).
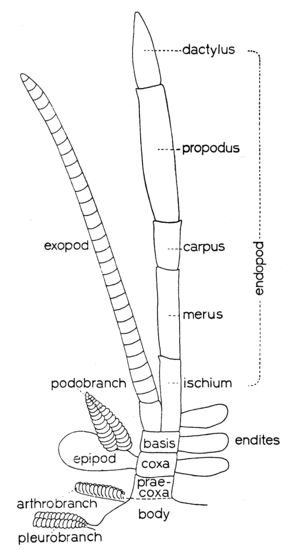
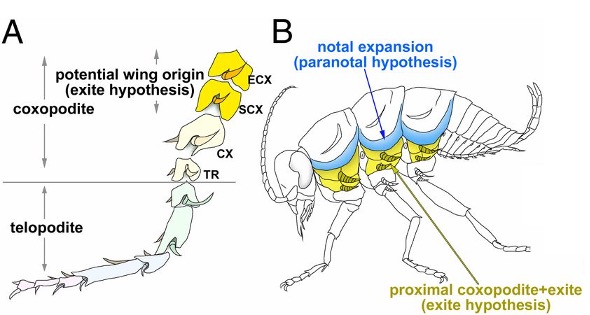
La première, proposée par la paléo-entomologiste tchèque Jarmila Kukalova-Peck, appelée théorie de l’exite, tient compte du fait que les insectes et les crustacés partagent un ancêtre commun. Les pattes de certains crustacés, composées d’un appendice ventral ou interne (endopodite) et d’un appendice dorsal ou externe (exopodite), sont dites « biramés ». Les insectes ne possédant qu’un appendice, les ailes dériveraient d’une excroissance (branchie par exemple) de l’exopodite des crustacés (figure 1). Dans ce modèle, la patte proximale aurait ainsi été incorporée dorsalement dans la paroi du corps (thorax), là où se situent les ailes actuellement.
La seconde, appelée théorie paranotale, propose que les ailes auraient évolué à partir de plaques de la paroi corporelle (lobes paranotaux), similaires à celles retrouvées chez les insectes aptères comme les lépismes argentés et qui sont absentes chez les crustacés (figure 2).
Malgré les preuves accumulées, aucun consensus scientifique en faveur de l’une ou de l’autre théorie n’a pu être établi, chacune des théories semblant aussi convaincante l’une que l’autre.
Récemment, des chercheurs ont proposé un modèle à double origine dans lequel les ailes résulteraient de la fusion de la paroi corporelle (thorax) et de la patte (exopodite). Pour déterminer ce processus évolutif, il est nécessaire d’identifier les structures impliquées et homologues en comparant l’anatomie des crustacés et celle des insectes.
Deux scientifiques, Heather S. Bruce et Nipam H. Patel (Université de Berkeley, Californie, et Université de Chicago), ont étudié le développement embryonnaire de Parhyale hawaiensis, un crustacé amphipode (plusieurs paires de pattes antérieures spécialisées dans le déplacement sur le sol et la préhension de la nourriture, et plusieurs paires de pattes postérieures servant à la nage) (lien).
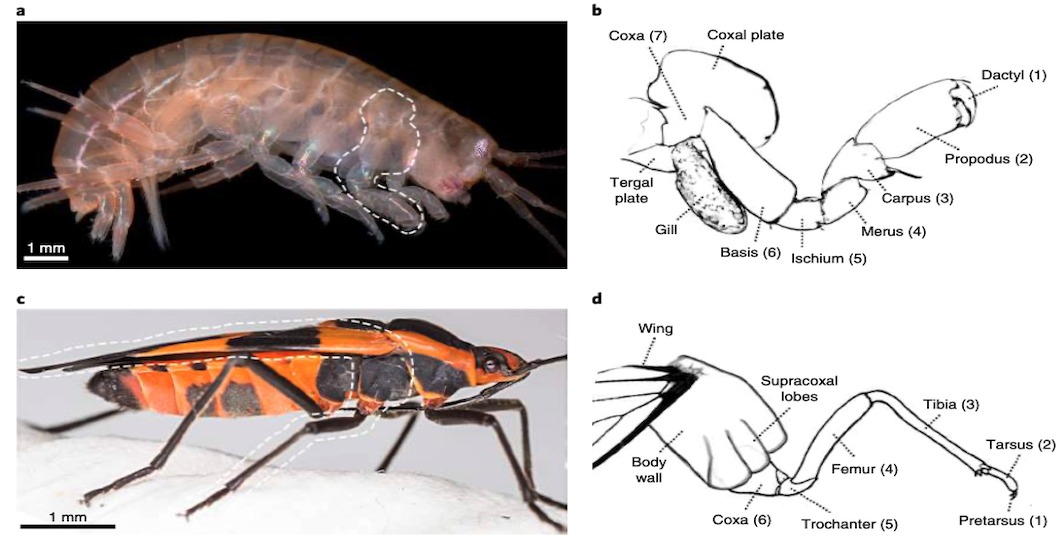
Cette espèce a la particularité de posséder des pattes dites « uniramées » (i.e. ne comportant qu’une seule partie par opposition aux pattes « biramées ») où chaque segment est clairement identifiable.
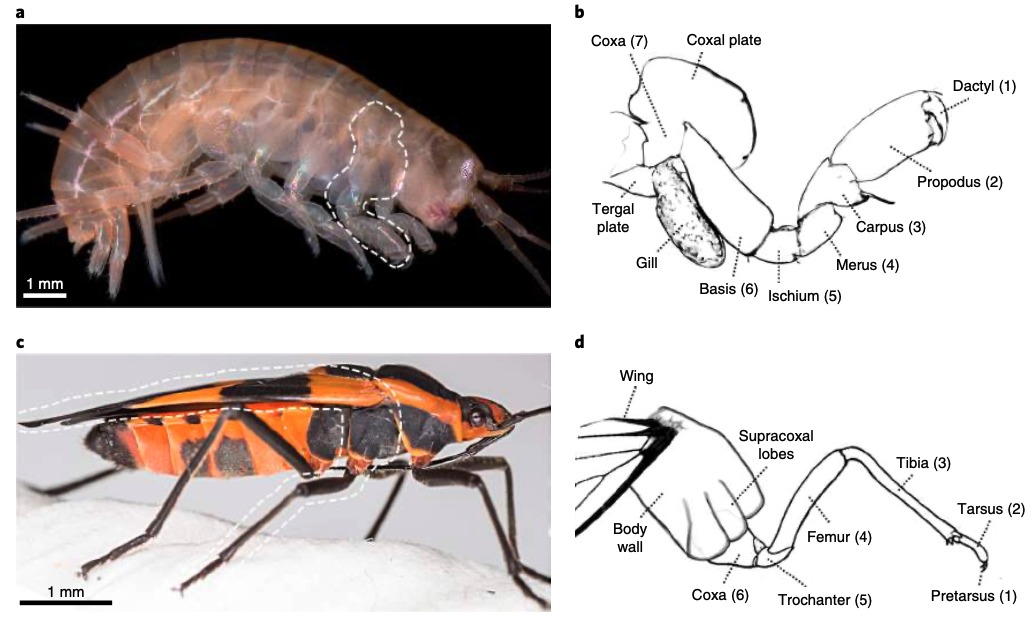
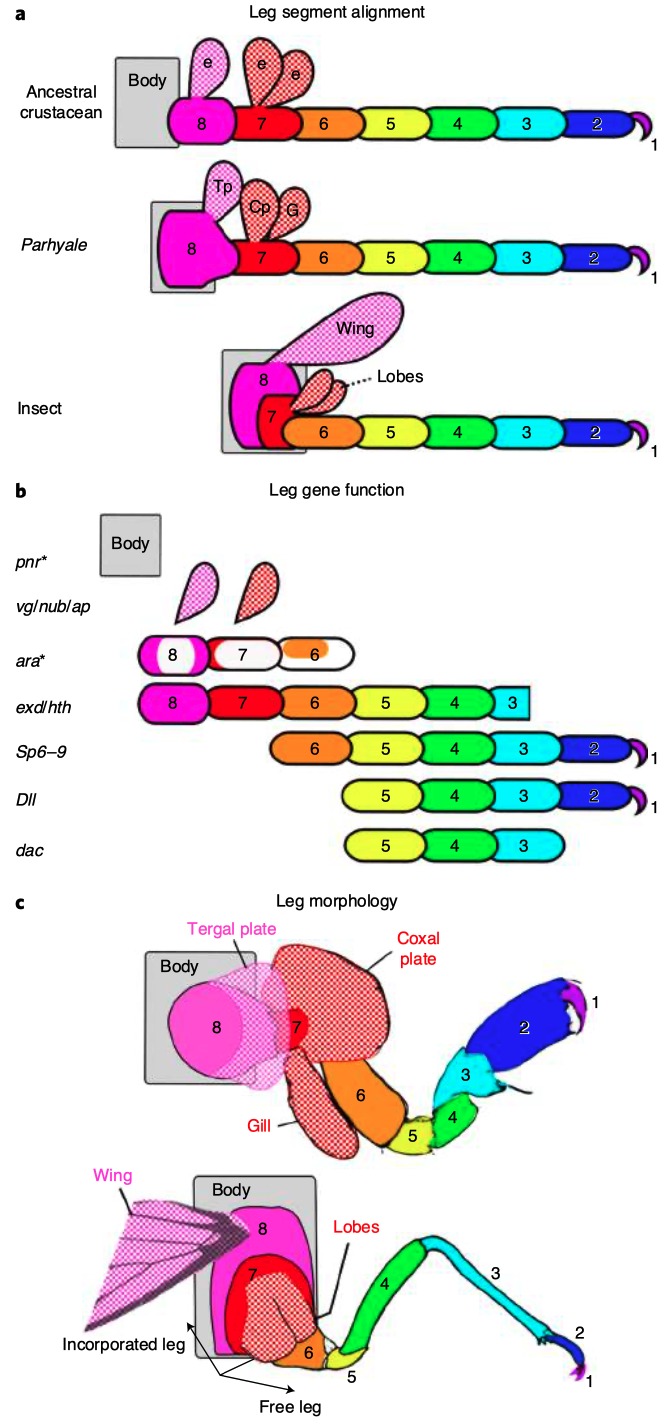
Chez un grand nombre de crustacés, les pattes sont constituées de 7 segments, tandis que chez les insectes, les pattes en ont uniquement 6 (figure 3 ci-dessous).
Ces chercheurs ont étudié le développement embryonnaire des pattes ainsi que l’expression de cinq gènes associés (méthode CRISPR-Cas9) : distaless (DII), Sp6-9, teckel (dac), extradenticle (exd) et homothorax (hth). Ces gènes sont également connus et étudiés chez trois espèces d’insectes dont la drosophile pour leur intervention dans le développement des différents segments des pattes.
Un lien entre pattes et ailes se confirme !
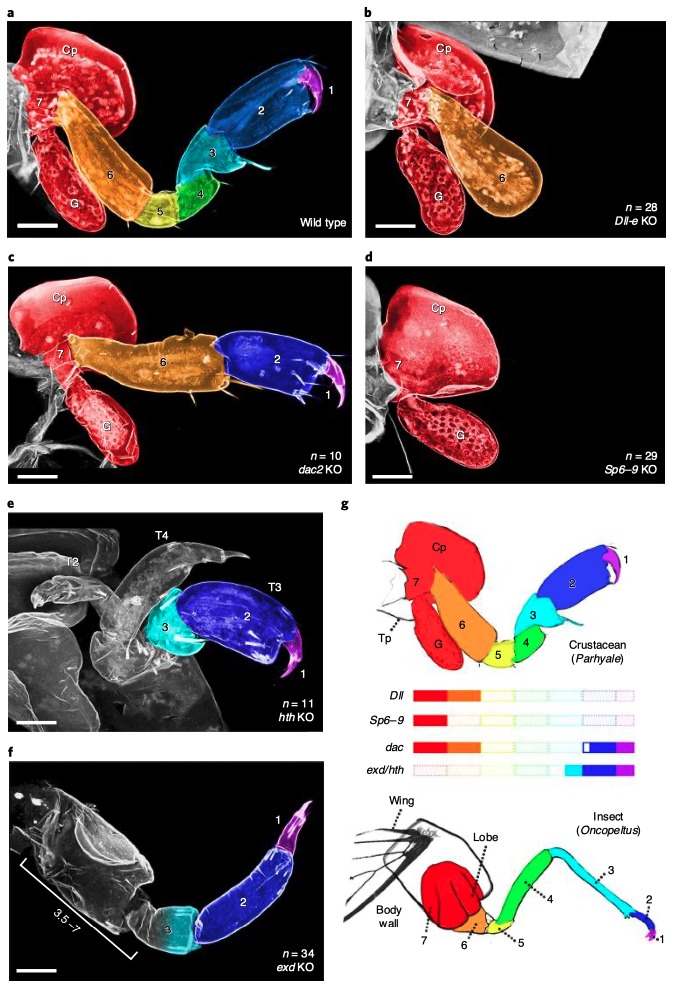
Les premiers résultats des expressions géniques ont mis en évidence une correspondance entre les structures des pattes de l’insecte et celles du crustacé (figure 4 ci-dessous).
Les expressions des gènes DII, Sp6-9, dac, exd et hth sont similaires dans la mise en place des différents segments chez l’insecte et le crustacé avec pour correspondance : Dactyl/Pretarses ; Propodus/Tarses ; Carpus/Tibia et ainsi de suite (figures 3b et 3d). Fait intéressant, chez Parhyale, un segment de la patte n’est pas présent chez l’insecte : le « coxa crustacé ». Ce segment ancestral pourrait avoir simplement disparu chez les insectes, ou, d’après la théorie de l’exite, il aurait été incorporé dans la paroi corporelle de l’insecte.
Ainsi, la paroi corporelle des insectes serait constituée de deux régions distinctes : une région dorso-médiane correspondante à la paroi du thorax (non homologue à la patte) et une région ventrale formée des segments proximaux de la patte (figure 6).
De manière plus précise :
- Le gène DII est nécessaire chez le crustacé et la drosophile pour le développement des segments 1 à 5 décomptés à partir de l’extrémité distale (figure 4b)
- Le gène dac, le crustacé exprime deux versions paralogues (deux versions provenant d’une version ancestrale) : dac1 qui ne semble pas avoir de phénotype visible associé, et dac2 intervenant dans la mise en place des segments 3 à 5 (figure 4c). Chez la drosophile, le gène dac intervient quant à lui dans la mise en place du trochanter (segment 5)
- Le gène Sp6-9 intervient pour le développement des segments 1 à 6 de manière similaire chez Parhyale et la drosophile (figure 4d).
- Les gènes Exd et hth interviennent quant à eux dans la formation des segments 4 à 7 à la fois chez le crustacé et la drosophile. Leur suppression entraine des disparitions ou des fusions de segments (figures 4e et 4f)
Pour tester l’hypothèse de l’incorporation de la coxa ancestrale des crustacés dans la paroi corporelle des insectes, les scientifiques ont recherché des gènes associés à ce processus. L’idée repose sur le fait que les gènes exprimés dans la zone dorsale chez la drosophile, correspondant à la vraie paroi corporelle, devraient être exprimés dans le tissu le plus dorsal de Parhyale. Inversement, les gènes exprimés dans la paroi latérale gauche et droite du corps de la drosophile, mais pas dans l’aile ou la patte, pourraient être exprimés dans le coxa de Parhyale.
Deux gènes ont ainsi été identifiés : pannier (pnr) qui est exprimé chez la drosophile sur une seule bande dorsale comme cela était suggéré, et les gènes du complexe Iroquois, exprimés chez la drosophile dans la paroi latérale du corps (formant un C autour de l’aile) et qui semblent nécessaires pour édifier la paroi du corps autour de l’aile.
La drosophile étant un insecte holométabole (métamorphose complète – lien), l’embryogenèse de la larve ne permet pas de suivre le développement des pattes. Pour remédier à cela, les scientifiques ont donc utilisé comme modèle d’étude Tribolium castaneum (Tenebrionidae – Coleoptera), dont la larve possède des pattes, pour réaliser les comparaisons avec Parhyale.
L’expression du gène pnr est similaire chez les deux espèces au point qu’elle peut être superposée (figure 5). Il s’exprime dans les tissus les plus dorsaux correspondant à une véritable structure de la paroi corporelle. L’étude du développement embryonnaire montre que la coxa de l’insecte dérive d’une intégration de la coxa du crustacé. Ainsi, chez Tribolium, la coxa s’aplatit pour former un élément de la paroi latérale du corps. Ces résultats apportent la preuve que la coxa du crustacé ancestral s’est insérée dans la paroi corporelle de l’insecte.
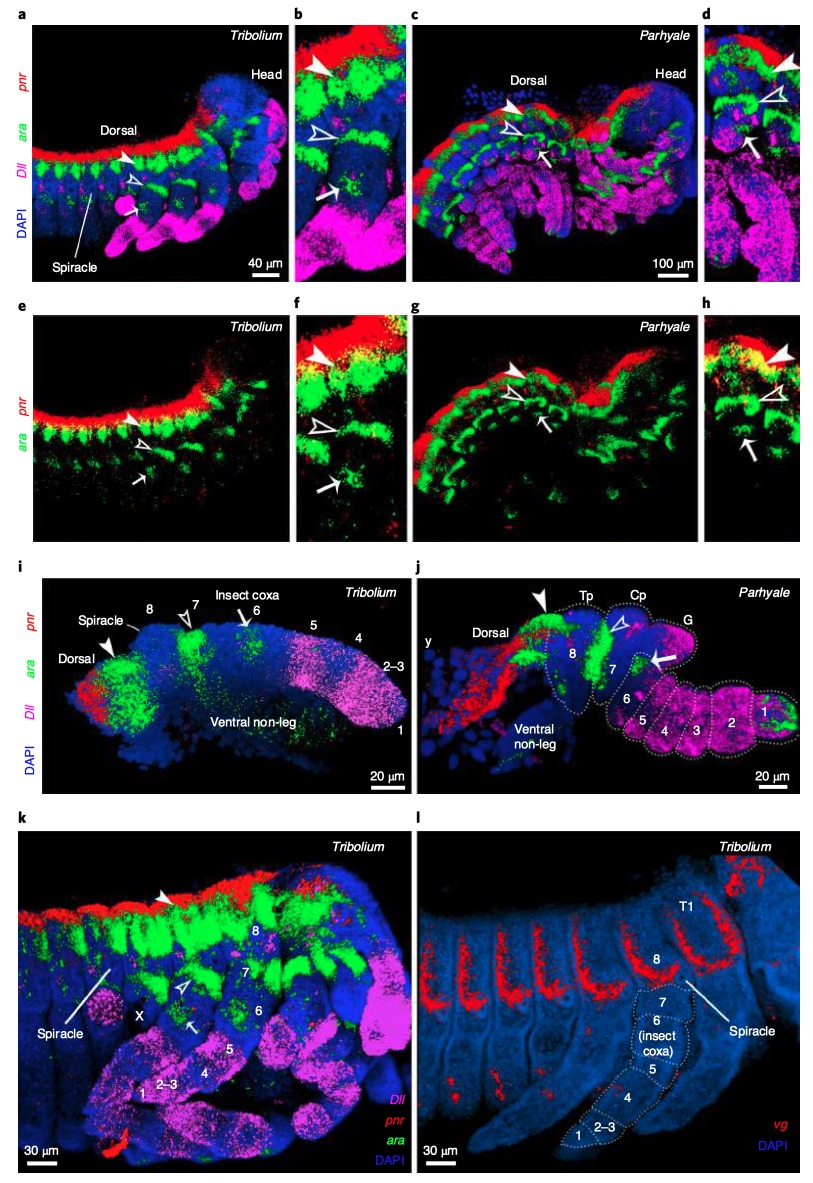
De plus, l’expression de ces gènes, encadrant la région de l’aile présumée chez Tribolium et la plaque tergale (plaque située à la base ventrale de la patte – figure 2) chez Parhyale, suggèrent que l’aile de l’insecte peut être homologue à la plaque tergale des crustacés (figure 6a).
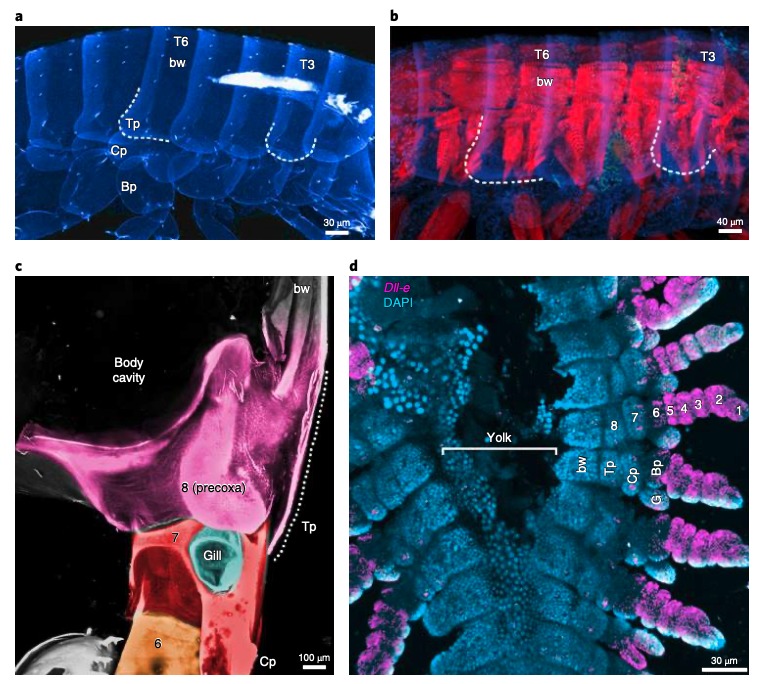
Chez Parhyale, la plaque tergale et les deux excroissances externes fixées à la base de la patte (les branchies et la plaque coxale) expriment et nécessitent les mêmes gènes que ceux intervenant dans le développement des ailes d’insectes. La plaque tergale, fixée au thorax (paroi corporelle) est singulière car, traditionnellement, elle est considérée comme associée aux segments de la patte. Les scientifiques ont donc formulé l’hypothèse qu’une partie de la paroi corporelle de Parhyale dériverait d’une intégration d’un segment de la patte comme chez les insectes.
De nombreux crustacés possèdent un segment supplémentaire de patte : la précoxa. Pour déterminer si Parhyale possède un vestige de précoxa ayant été incorporé dans la paroi corporelle, ils ont examiné les embryons, les juvéniles, et les adultes de Parhyale par microscopie confocale (microscope optique ayant la propriété de réaliser des images de très faible profondeur de champ, environ 400nm). Ainsi, il a été identifié une structure précoxale répondant aux critères d’un segment de patte visible de la paroi du corps (deux fois la longueur du coxa) (figure 6c). Cette structure forme une articulation musculeuse s’étendant à un autre segment de la patte (figure 6). De plus, chez les embryons de Parhyale, huit subdivisions peuvent être observées dans les pattes en développement (figure 6d) Ainsi, une grande partie de la paroi latérale du corps de Parhyale semble dériver d’un segment de patte proximale ancestral, comme chez les insectes.
Finalement, pourquoi cette intégration ?
Si les pattes des crustacés comportent deux segments supplémentaires de plus que celle des insectes : précoxa (segment 8) et coxa des crustacés (segment 7) (figure 2), la question est : que sont-ils devenus chez ces derniers ?
Les données provenant de l’expression des gènes suggèrent que ces deux segments de pattes ancestraux ont étés intégrés dans la paroi thoracique de l’insecte et que l’aile de l’insecte est homologue à la plaque tergale chez Parhyale. Il semblerait donc que les ailes des insectes auraient évolué à partir d’une structure existante chez des crustacés ancestraux (figure 7).
Cette théorie fut pour la première fois présentée par le zoologiste danois Hans Jacob Hansen (1855-1936) en 1893, puis par de nombreux scientifiques comme le zoologiste et entomologiste allemand Richard Heymons (1867-1943) en 1899 ou encore l’arachnologiste américain Henry Elisworth Ewing (1883-1951) en 1928 et l’entomologiste anglais Augustus Daniel Imms (1880-1949) en 1937. De nombreuses observations embryologiques, morphologiques, moléculaires et phylogénétiques sont depuis venues conforter ce modèle.
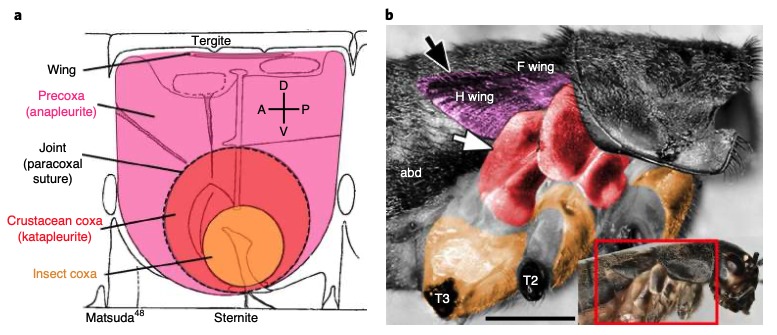
En effet, l’étude de nombreux embryons d’insectes a mis en évidence la fusion des segments proximaux des pattes dans la paroi corporelle. Chez les adultes, la structuration des plaques du thorax entourant la patte évoque deux segments de patte aplatis séparés par une articulation avec des insertions musculaires (figure 7). Ce reste d’articulation peut également s’observer au niveau moléculaire : le gène Serrate, qui marque les articulations du segment de la patte, est également exprimé à la frontière entre les deux segments des patte incorporées dans la paroi du corps chez Tribolium.
Bien que l’aile provienne d’une excroissance présente sur le segment 8 de la patte chez les insectes, il semblerait que pour certains d’entre eux, comme les Orthoptères, les Hémiptères et les Coléoptères, ce soit à partir du segment 7 (lobe supra-coxal). Deux scientifiques japonais, Mashimo Y. et Machida R., ont décrit en 2017 le développement embryonnaire du sous-coxa du grillon (segment 7), à partir duquel on observe le développement du lobe. Notamment, les gènes wing et exite sans ailes et vestigiaux sont exprimés non seulement dans le tissu présumé de l’aile mais également là où le lobe se formerait chez le Ténébrion meunier (Tenebrio castaneum), le grillon (Gryllus assimilis), la mouche des fruits (Drosophila melanogaster).
De récentes mises à jour de la phylogénie des crustacés permet de retracer l’histoire évolutive du segment 8 : transformation de la patte de crustacés en ailes d’insectes. Par exemple, la plaque tergale de Parhyale ressemble aux lobes pleuraux des crustacés céphalocarides et aux lobes paranotaux d’hexapodes sans ailes comme le lépisme argenté.
D’un point de vue moléculaire, des études ont confirmé l’expression de gènes au niveau de segments thoraciques dorsaux intervenant dans la formation des ailes chez un insecte aptères comme P. unimaculatus.
Enfin, l’incorporation de segments de la patte proximale dans la paroi corporelle n’est pas rare chez les arthropodes. Les mille-pattes et les myriapodes semblent également avoir incorporés un segment de la patte proximale dans la paroi corporelle. Les crustacés isopodes terrestres ont fusionné non pas un mais deux segments de la patte proximale dans le corps (précoxa et coxa), de sorte que la base (segment 6) constitue maintenant le premier segment de la patte libre. Notamment, ce sont les deux mêmes segments de patte proximale qui sont supposés avoir été incorporés dans la paroi corporelle de l’insecte.
Il est intéressant de noter que, pour tous ces groupes, l’incorporation de segments de pattes proximales dans le thorax résulterait de la nécessité de soutenir le poids de l’animal subissant la gravité (adaptation à la sortie des eaux : passage d’un milieu aquatique à terrestre). Cette intégration aurait ainsi permis de renforcer la patte pour la rendre plus résistante à la pression.
Synthèse des homologies de structures entre crustacés ancestraux et insectes, et des processus évolutifs d’intégration des segments et de la formation des ailes chez les insectes
Bibliographie
- Bruce H.S & Patel N. H. (2020) : Knockout of crustacean leg patterning genes suggests that insects wings and body walls evolved from ancien leg segments. Nature Ecology & Evolution, (4)1703-1712 (lien)
- Clark-Hachtel C. M. ; Linz D.M. & Tomoyasu Y. (2013) : Insights into insect wing origin provided by functional analysis of vestigial in the red flour beetle, Tribolium castaneum. PNAS, 110(42),16951-16956 (lien)
Ouvrages sur cette thématique
- The Insects : Structure & Function (5th edition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
- Evolution of the Insects (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
- The Insects: An Outline of Entomology (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
- The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
- Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)