Platerodrilus : cas de néoténie chez les insectes
Par Benoît GILLES
Au cœur des forêts tropicales et subtropicales de l’Asie du sud-est, de l’Himalaya et de la Chine jusqu’à Java, Bornéo et les Philippines, évoluent d’étranges et discrètes créatures qui pourraient sembler tout droit sorties du Paléozoïque (-540 Ma ; – 250Ma).
Possédant un corps aplati et élargi similaire à celui des trilobites (figure 1 – vidéo en bas de page) dont elles tirent leur surnom en anglais « trilobite beetles », ces créatures sont en fait des insectes appartenant à la famille des Lycidae et à la tribu des Platerodrilini. Bien que ces coléoptères soient connus depuis le milieu du 19ème siècle et abondants dans de nombreuses régions, notamment ceux du genre Platerodrilus qui est le plus diversifié et répandu, leur écologie et leur biologie demeurent en grande partie inconnues.

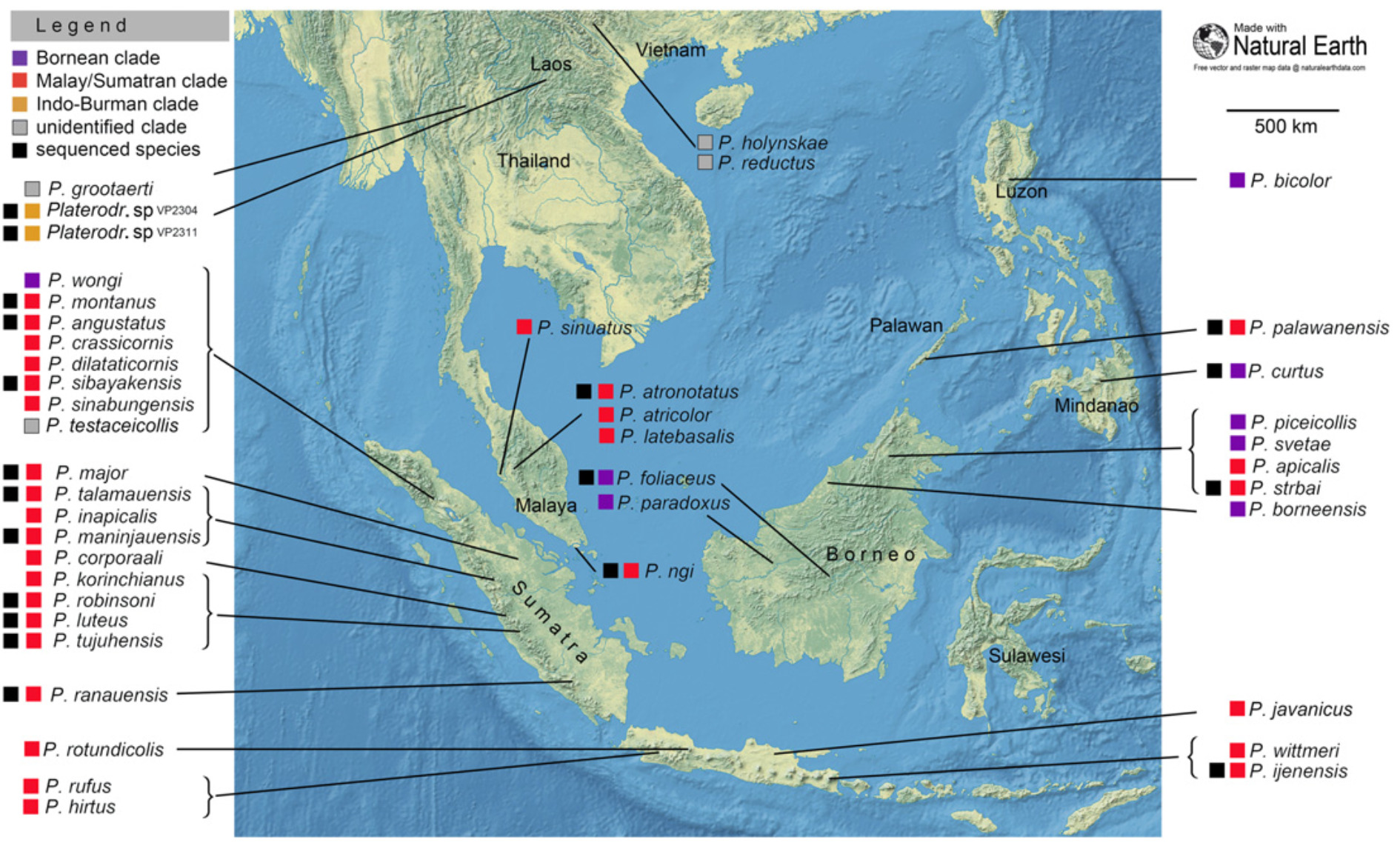
Le genre Platerodrilus est constitué de 42 espèces réparties en trois zones géographiques : Indo-Birmanie, Sundaland (Malaisie, Bornéo, Sumatra et Java) et Philippines (comprenant l’archipel de Palawan (figure 2).
Cas rare de néoténie

La néoténie caractérise des organismes vivants qui conservent des caractères juvéniles ou larvaires à l’état d’adulte sexué. L’exemple le plus connu chez les vertébrés est l’Axolotl (Ambystoma maxicanum), une salamandre dont les adultes, essentiellement aquatiques, ressemblent à des têtards avec leurs branchies (figure 3).
Les coléoptères néoténiques Lycidae constituent quant à eux un cas de néoténie chez les insectes : seules quelque 100 espèces de Lycidae sont néoténiques, soit environ 3% des 4 300 espèces décrites.
De manière surprenante, la néoténie concerne chez ces insectes uniquement les femelles. Vivant plusieurs années, elles demeurent toute leur vie sous une forme « larvaire » aptère (adulte larviforme), ne différant de la larve que par quelques traits morphologiques (paragraphe suivant). Les mâles, ailés, réalisent, quant à eux, une métamorphose complète (holométabolie) typique des coléoptères.
Nous sommes ainsi face un exemple rare où femelles et mâles ne possèdent pas les mêmes stades de développement.
Remarque : le passage de larves mâles en adulte n’a pu être documenté qu’a de rare fois au sein d’élevages, par exemple par l’entomologiste Mjoberg en Malaisie et par l’entomologiste Wong à Singapour (Mjöberg, 1925 ; Wong, 1996).
Ces insectes sont utilisés comme groupe modèle lors d’études en génétique évolutive et pour l’étude des fluctuations climatiques (zoogéographie) par exemple (paragraphe suivant sur phylogénie).
Morphologie
Autre élément singulier, il existe un important dimorphisme sexuel.
Les femelles mesurent de 3 à 7 cm alors que les mâles mesurent quant à eux entre 6 et 10 mm, voire 1,3 mm chez certaines espèces du genre Alyculus. Bien que les femelles soient inconnues pour la plupart des espèces et des genres, ce dimorphisme a été constaté au sein de plusieurs genres comme Platerodrilus, Macrolibnetis et Lyropaeus (figure 4).

- Description morphologique d’une larve femelle Platerodrilus svetae
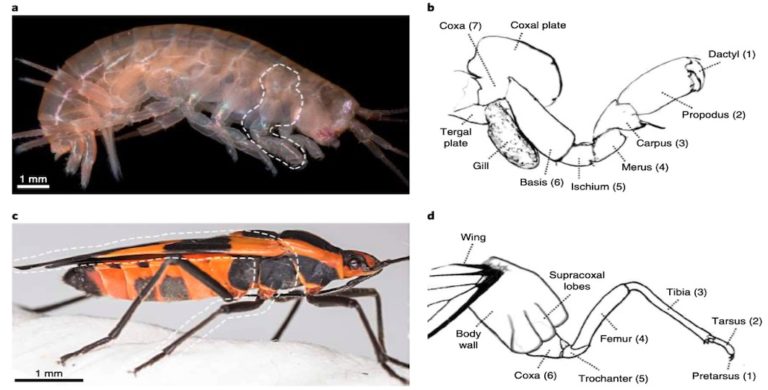
Le corps est composé de trois parties distinctes : tête, thorax, abdomen. Allongé et fortement sclérifié, il est globalement de couleur brun foncé avec les extrémités des excroissances de couleur orangé à jaune en fonction des espèces (figure 5). La tête, les côtes thoraciques, les appendices abdominaux (pleures) et les pattes sont recouverts d’une cuticule épaisse – lien). La partie antérieur (pronotum) est plus large au niveau des segments thoraciques qu’au niveau de l’abdomen.
La cuticule arbore des aréoles (surfaces circulaires) polygonales fermées, fortement pigmentée et portant de nombreuses sensilles (lien), en moyenne 340 par mm2.
La tête, de très petite taille par rapport à celle du prothorax, est plus longue que large, complétement rétractable dans le protothorax. La capsule céphalique ouverte ventralement est composée de quatre sous-unités : une plaque dorsale avec trois paires de soies longues, une paire de plaques latérales avec chacune deux longues soies, une plaque ventrale semi-circulaire avec une paire de soie et une paire d’apophyses postérodorsales étroites (figures 4 et 5).

Les antennes, également rétractables, sont constituées de deux segments, un antennomère annuliforme (forme d’un anneau) étroit, un antennomère grand et robuste multilobé à son extrémité membraneuse (figure 6).
Les pièces buccales sont composées de mandibules, de maxilles portant des palpes maxillaires à 3 segments se rétrécissant apicalement et de palpes labiaux à deux segments (figure 6) (lien).

Les tergites thoraciques, considérablement plus larges que les tergites abdominaux, sont grossièrement ponctués. Le prothorax arbore une forme trapézoïdale et une paire de tubercules dans sa partie proximale. Le méso- et le métathorax, rectangulaires, en possèdent quant à eux deux paires chacun (figure 5).
Les pattes, composées de cinq segments, possèdent ventralement une brosse de poils courts et denses. La coxa est libre et allongée, le trochanter est divisé en deux parties par une ligne membraneuse portant des sensilles espacées (figure 7).

Chaque tergite abdominal possède une paire d’épipleures (excroissances sur la face dorsale) proéminents dirigés vers l’arrière et une paire d’hypopleures (excroissance en face ventrale) plus petits également dirigés vers l’arrière (une paire étant également présente sur chaque sternite) (figure 5).
- Description morphologique d’une femelle adulte larviforme de Platerodrilus svetae
La femelle larviforme est morphologiquement similaire à la forme larvaire au dernier stade.
Les principales différences entre ces stades se situent au niveau de la tête, où les mandibules sont davantage écartées l’une de l’autre, plus épaisses et uniformes et de la surface de la cuticule qui est moins pigmentée, plus fine avec de nombreuses aréoles munies d’épines de différentes tailles et des sensilles en moindre quantité (1 à 2 par mm2) (figure 8).
Le système circulatoire est composé d’une aorte centrale et d’un coeur tubulaire dans chaque segment abdominal (figure 8)
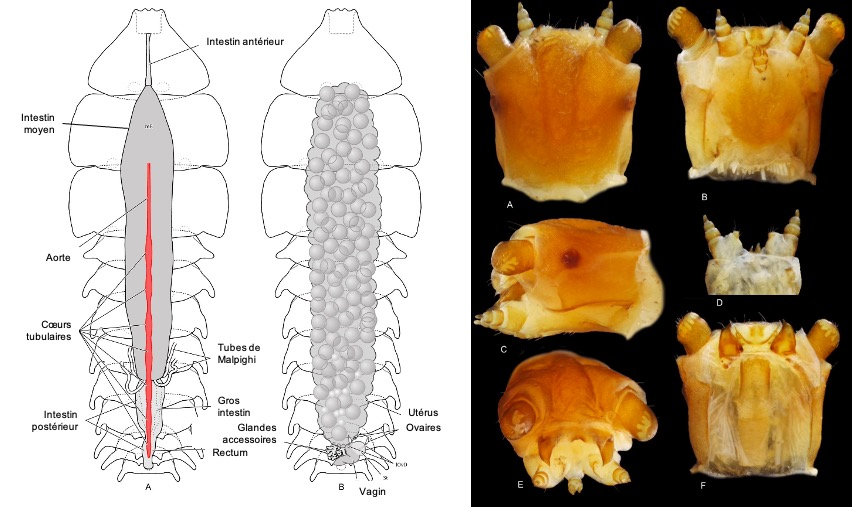
Le système digestif de la femelle larviforme est similaire à celui de la forme larvaire. Le tube digestif, simple, comprend notamment un intestin antérieur court, légèrement élargi dans sa partie distale, un intestin moyen tubulaire non segmenté, significativement déformé par un utérus hypertrophié. Chez la larve, l’intestin moyen est plus large et connecté à un réseau dense de trachées (figure 8).
A l’interface entre les intestins moyen et postérieur s’insèrent deux paires de tubes de Malpighi chez la femelle larviforme: ils sont, semble-t-il, plus nombreux chez la larve (figure 8).
L’intestin postérieur est court chez la femelle larviforme et beaucoup plus long et formant une boucle entre les segments IV et VI de l’abdomen chez la larve. Chez les deux formes, l’intestin postérieur est différencié en section plus large avec une surface plissée (figure 8).
Le système nerveux est similaire entre la forme adulte et la forme larvaire à deux détails près : chez la larve, le ganglion métathoracique est sensiblement plus gros que les ganglions précédents.
Les œufs sont sphériques et de couleur jaune à blanchâtre. Avec 1 mm de diamètre, ils sont particulièrement gros par rapport à la taille de l’insecte (figure 9).

- Les mâles
Bien qu’ailés, ils n’ont la capacité de voler que sur de courtes distances dans les strates basses des forêts tropicales humides à la recherche de femelles. Ils ne vivent que très peu de temps à l’état adulte (figure 4).
Ecologie
Ces Coléoptères se rencontrent uniquement dans des environnements tropicaux humides car les femelles larviformes s’alimentent essentiellement de nourriture liquide en raison de pièces buccales de petite taille et non adaptées à broyer la matière organique.
La fécondité des femelles est plutôt limitée. Elles pondent des œufs de grande taille d’où éclosent des larves de taille comparable à celle des mâles adultes !
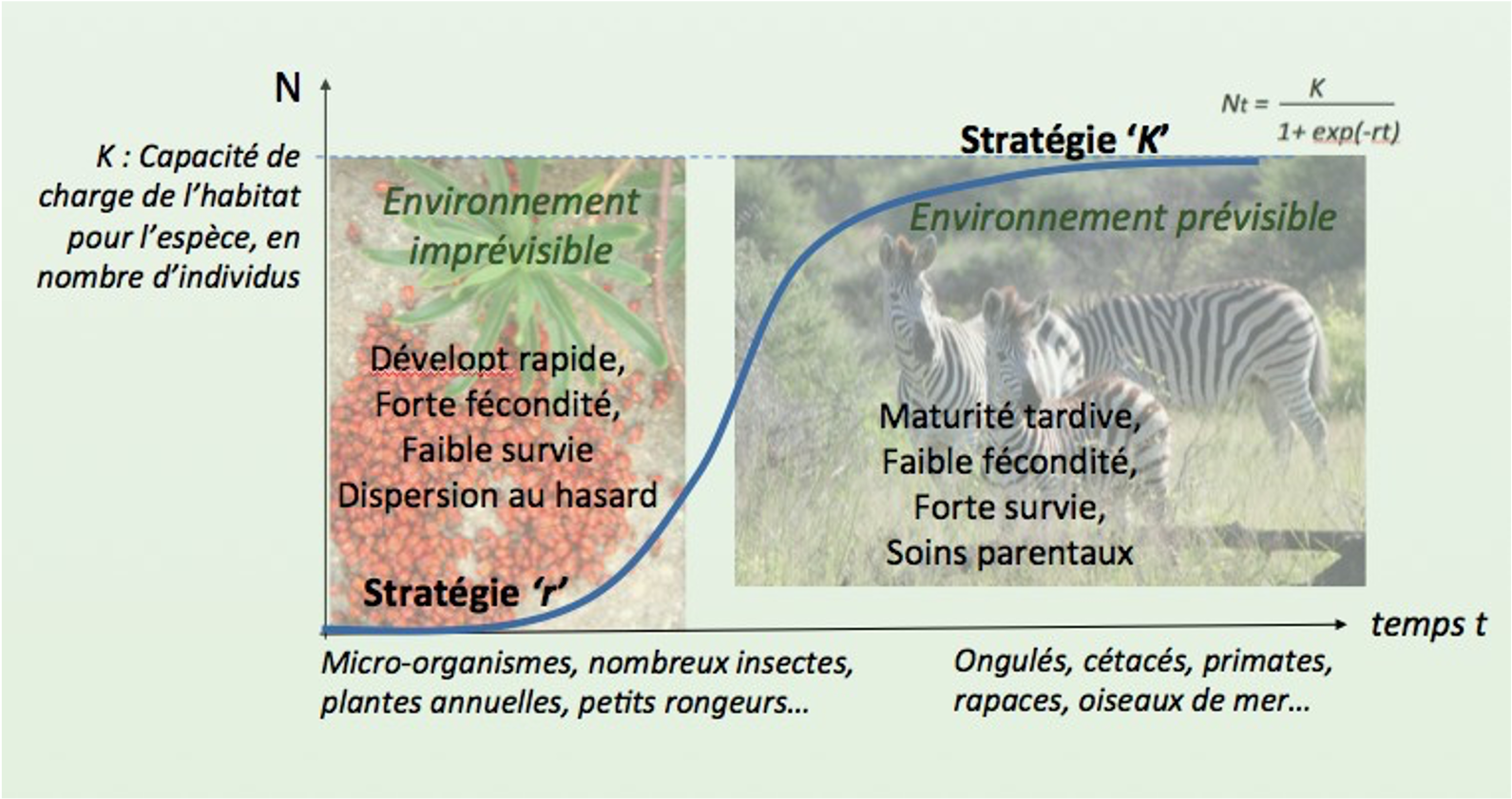
Une telle stratégie de reproduction est coûteuse en énergies et est considérée comme un exemple de stratégie K.
Les mâles, au contraire, investissent un minimum d’énergie dans la reproduction par rapport au femelle.
La stratégie K est associée à des conditions de vie prévisibles et des risques de mortalité faibles où les espèces peuvent allouer des ressources et investir dans la survie de la progéniture, peu nombreuse : c’est le cas d’un grand nombre de vertébrés comme les éléphants, les baleines, les oiseaux. A contrario, la stratégie r consiste à pallier des environnements instables ou des risques élevés de mortalité de la descendance par une forte fécondité comme pour les poissons, les batraciens, les crustacés ou les micro-mammifères (figure 10).
Phylogénie
Depuis les premières descriptions du début du 20ème siècle, déterminer la place taxonomique des Lycidae néoténiques à partir de traits morphologiques n’est pas chose aisée sachant que pour un grand nombre d’espèces les couples ne sont pas connus (quelques accouplements seulement observés au sein du genre Platerodrilus – (voir notamment Mjöberg 1925, Wong 1996), et que les critères morphologiques demeurent arbitraires.
Certaines études ont intégré les Platerodrilini au sein du clade Cantharidoidea (Lawrence et al., 2011), et les rapprochent du complexe regroupant les familles des Cantharidae, Omethidae, Telegeusidae et Phengodidae (Branham & Wenzel, 2003).
D’autres études placent les Platerodrilus dans le clade Elateroidea mais elles restent toutefois incompatibles avec celles produites à partir d’analyses moléculaires (Bocak & Bocakova, 1990, 2008 ; Branham & Wenzel, 2003 ; Lawrence et al., 2011 ; Kundrata et al., 2014 ; Bocak et al., 2016 ; Masek et al., 2018 ; Zhang et al., 2018) (figure 11).
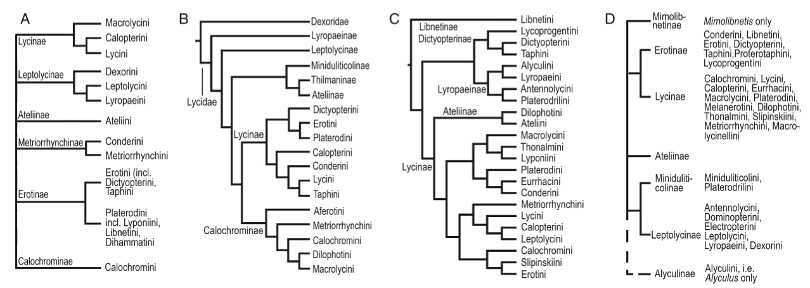
Après trois décennies de débats, l’histoire évolutive de ces insectes demeure non résolue. Cependant, des analyses moléculaires récentes ont apporté de nouveaux éléments.
L’équipe menée par Ladislav Bocak de (Université d’Olomouc – Tchéquie) (2019) a exploité des données à la fois génomiques et transcriptomiques (liées aux ARN), fournissant des informations phylogénétiques indépendantes de la morphologie.
Leurs résultats répondent à trois questions précédemment controversées : (i) la classification au niveau de la tribu et de la sous-famille ; (ii) l’apparition multiple de la néoténie au sein du clade et la convergence des traits morphologiques partagés chez les mâles des espèces aux femelles néoténiques ; (iii) l’évolution des traits morphologiques, notamment ceux à fonction adaptative, associés au mimétisme ou affectés par la sélection sexuelle.
Le clade est donc constitué de trois taxons monophylétiques distincts : Lycinae, Lyropaeinae et Metriorrhynchinae.
Il semblerait que la diversification au sein de ces sous-familles ait été rapide (branches courtes dans l’arbre phylogénétique)
La sous-famille des Lycinae se compose des tribus Conderini et de quatre tribus gondwaniennes : Leptolycini, Platerodini, Lycini et Calopterini. La sous-famille des Lyropaeinae intègre l’ensemble des espèces asiatiques à l’exception de la tribu des Ateliini, et la sous-famille des Metriorrhynchinae intègre les tribus Lycoprogenthini, Dihammatini, Dilophotini, Libnetini et Metriorrhynchini (figure 12).

Ces résultats mettent en évidence que la néoténie serait apparue de manière indépendante à plusieurs reprises dans les différents taxons.
Cette convergence morphologique des femelles et des mâles résulterait de processus évolutifs aboutissant à de l’homoplasie (acquisition indépendante de caractères similaires) : convergence évolutive.
Mécanismes génétiques et mimétiques
A l’instar des femelles, les mâles des espèces néoténiques présentent des caractères morphologiques similaires comme un corps de petite taille, un nombre réduit d’antennomères, des pièces buccales atrophiées ou réduites (mandibules rudimentaires, palpes maxillaires et labiales pointues distalement), des élytres raccourcis comme chez Mimolibnetis apicalis ou Alyculus sp., une perte ou une importante réduction des ailes comme chez Dexoris chome ou Cautires apterus, l’absence d’éperons tibiaux et des tarsomères minces avec des coussinets tarsiens réduits (figure 12).
Les mécanismes moléculaires n’étant pas compris et connus, il est pour l’heure impossible de faire la différence entre des traits morphologiques modifiés en raison d’une évolution adaptative ou résultant de la disparition de l’holométabolie chez les femelles (lien). Par exemple, la réduction des pièces buccales chez les mâles pourrait avoir suivi la réduction de leur espérance de vie et l’absence d’alimentation durant le stade adulte, ou la différenciation limitée de plusieurs parties du corps au cours de la métamorphose.
La réduction de la taille des élytres et des ailes pourrait être associée à la faible capacité de dispersion des femelles larviformes du fait de leur mobilité réduite. Les mâles auraient moins de distance à parcourir pour trouver une femelle.
Contrairement aux pièces buccales, l’hypothèse fondée sur la réduction de la durée de vie ou sur la distance à parcourir, ne peut pas, quant à elle, être appliquée aux modifications des tarsomères. Il est probable qu’une différenciation structurelle incomplète des appendices, notamment des pattes et des structures dérivées, serait engendrée par une reprogrammation ontogénique (développement de l’organisme) affectant les deux sexes, l’influence étant toutefois moins marquée chez les mâles.
Cependant, des exceptions existent. Les mâles d’espèces néoténiques apparentées présentent toutefois des traits morphologiques différents. Par exemple, ceux de certaines espèces du genre Platerodrilus possèdent des corps de grande taille et des pièces buccales non atrophiées, d’autres ont un corps large et des pièces buccales réduites et simplifiées comme le genre Lyropaeus,
À l’inverse, des taxons morphologiquement similaires peuvent ne pas être apparentés selon la phylogénie moléculaire. Par exemple, les Calopterini néoténiques placés morphologiquement parmi les Leptolycini, ne le sont plus à partir des analyses moléculaires.
La convergence d’adaptations et de traits morphologiques (homophasie) chez les femelles néoténiques est également favorisée par des pressions de sélection naturelle associées à des processus de mimétisme.
Comme chez les papillons Heliconius (lien), la sélection naturelle tendrait ici à faire converger toutes les espèces d’une même région, quel que soit leur proximité phylogénétique, vers un pattern (aspect morphologique) identique. Il s’agit d’un cas de mimétisme Müllérien (vs mimétisme Batésien). Ce phénomène semble amplifié par le fait que les femelles possèdent des couleurs aposématiques (couleurs dissuasives pour les prédateurs) à l’extrémité de leurs excroissances abdominales (épi- et hypopleures) (figures 1 & 13) et par la présence dans leurs tissus de molécules aversives dans leur corps dissuadant les prédateurs de les consommer (stratégie pour contrer leur faible capacité à se déplacer).
La convergence morphologique des espèces et le fait d’arborer des couleurs facilement repérables dans le sous-bois ombragé des forêts tropicales, renforce l’apprentissage d’un prédateur à ne pas consommer tel ou tel pattern. Un individu différent du pattern majoritaire dans une population sera rapidement contre-sélectionné.

Bibliographie
-
- Bocak L. ; Bocakova M. ; Hunt T. & Vogler A.P. (2008) : Multiple ancient origins of neoteny in Lycidae : consequences for ecology and macroevolution. Proceedings of the Royal Society, 275,2015-2023 (lien)
- BocakL. & Bocakova M. (2008) : Phylogeny and classification of the family Lycidae. Annales Zoologici, 58(4):695-720 (lien)
- Kusy D. et al. (2019) : Phylogenomic analysis resolves the relationships among net-winged beetles and reveals the parallel evolution of morphological traits. Systematic Entomology (lien)
- Makarov K.V. & Kazantsev S.V. (2022) : Larva, pupa or adult ? The female of Platerodrilus case. Diversity, 14,1063 (lien)
- Masek M. & Bocak L. (2014) : The taxonomy and diversity of Platerodrilus inferred from molecular data and morphology of adults and larvae. ZooKeys, 426:29-63 (lien)
- Masek M. ; Palata V. ; Bray T. & Bocak L. (2015) : Molecular phylogeny reveals high diversity, geographic structure and limited ranges in neotenic net-winged beetles Platerodrilus. PLOS ONE, 10(4) (lien)