Origine de la métamorphose chez les insectes
Par Benoît GILLES
La métamorphose est l’une des stratégies de vie les plus répandues chez les animaux. Les fortes différences morphologiques et physiologiques entre les formes larvaires et adultes impliquent l’exploitation d’habitats et de ressources alimentaires différentes, induisant des adaptations extrêmes pour des fonctions spécifiques telle que la dispersion.
Chez les amphibiens et de nombreux invertébrés marins, la métamorphose constitue une phase ancestrale du cycle de développement, trouvant son origine dans les profondeurs de l’histoire évolutive de ces groupes. A l’inverse, chez les insectes, la métamorphose est apparue au cours de leur évolution.
Les insectes arborent trois types de développement :
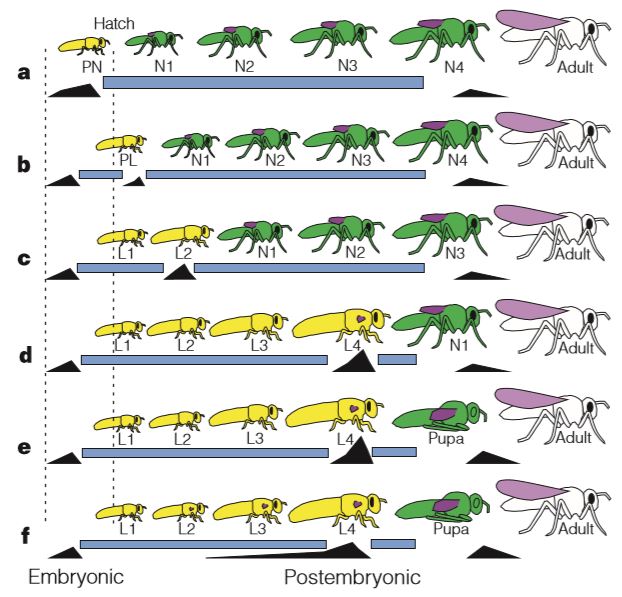
- Amétabole : développement se déroulant sans métamorphose, constituant la forme la plus ancienne. Aujourd’hui, ce groupe est représenté par les Ordres des Archéognathes (anciennement Thysanoures) et la famille des Lepismatidae (Ordre des Zygentomes), également appelés « poisson d’argent » ou lépisme. Le développement des amétaboles se composent de plusieurs mues successives où les jeunes sont semblables aux adultes, excepté les organes sexuels et génitaux qui apparaissent en fin de développement.
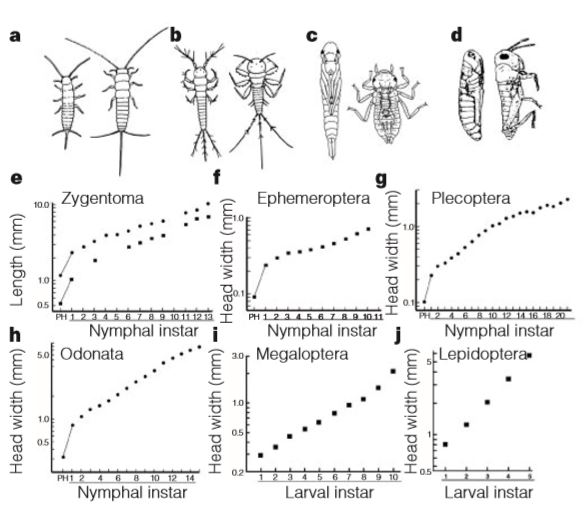
- Hémimétabole : également appelée métamorphose incomplète, elle représente une forme intermédiaire. Les jeunes, aussi nommés juvéniles, sont similaires aux adultes hormis les espèces ailées qui possèdent des bourgeons alaires repliés sur le dos, se transformant au stade adulte en ailes articulées fonctionnelles. Les hémimétaboles constituent un groupe paraphylétique (pas d’ancêtre commun vs monophylétique) incluant par exemple les Odonates (libellules), les Orthoptères (grillons, sauterelles, criquets), les Blattodea (blattes et termites), les Mantodea (mantes) et les Hétéroptères (punaises, pucerons, cigales…).
- Holométabole : désignant les insectes réalisant une métamorphose dite complète, les jeunes, appelés également larves, sont totalement différents de l’adulte tant morphologiquement que physiologiquement. La métamorphose se déroule durant une phase spécifique à l’intérieur d’une pupe (nymphe ou chrysalide) d’où émergera l’adulte. L’holométabolie apparaît durant le Permien (-300 à -250Ma) et constitue un groupe monophylétique (un même ancêtre commun vs paraphylétique) composé des Diptères (mouches), des Coléoptères, des Lépidoptères (papillons) et des Hyménoptères (abeilles, fourmis, guêpes).
La question que se posent les entomologistes est : « comment un cycle de développement composé d’une phase de métamorphose de type holométabole a pu apparaître au cours de l’évolution à partir d’une forme amétabole ? ».
Au début des années 2000, deux hypothèses étaient proposées.
L’entomologiste italien Antonio Berlese (1863-1927), remarquant la similarité entre les différentes formes larvaires et la morphologie de transition observée durant l’embryogenèse chez les hémimétaboles, proposa dès 1913 que les larves holométaboles seraient apparues par un processus de « dé-embryonisation » rendant l’embryon libre. L’éclosion prématurée, causée par une réduction de la quantité de vitellus (jaune d’œuf) stocké dans l’œuf, aurait engendré l’émergence d’une larve capable de se nourrir de manière autonome. La nymphe serait donc réduite au stade pupe
La seconde hypothèse, la plus largement établie, suggère que larves et nymphes sont similaires, le stade pupe se développant au fur et à mesure que la disparité entre les deux phases s’accentue.
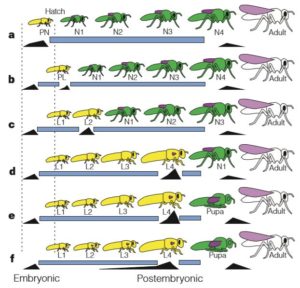
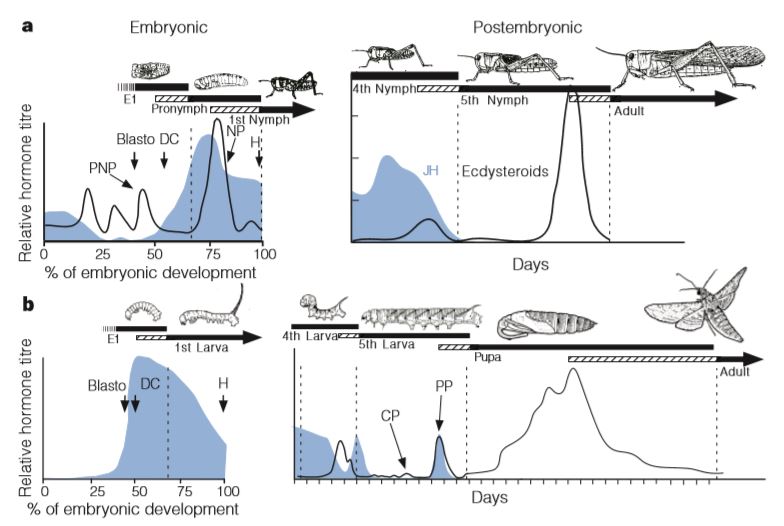
En 1999, les entomologistes américains James W. Truman et Lynn M. Riddiford (Département de Zoologie – Université de Washington) ont publié les résultats de leur étude portant sur le contrôle endocrinologique aux stades embryonnaire et post-embryonnaire. Ces derniers confortent la première hypothèse (Berlese) sur une origine embryonnaire de la métamorphose (figure 1 ci-dessous).
Forme et fonction du stade pronymphal
Chez de nombreux arthropodes comme les crustacés aquatiques et marins, la larve venant d’émerger de l’œuf (nauplius) présente une morphologie différente de celles des stades ultérieurs, souvent associée à un comportement de dispersion. Chez des arthropodes terrestres comme les isopodes (Crustacés), les scorpions et les mille-pattes (Myriapodes), le stade larvaire correspond à une phase où la larve ne peut ni se disperser, ni se nourrir. Pour survivre, elle bénéficie de soins maternels (protection) et survie grâce aux réserves vitellines. Cette période, dite pronymphale, correspond à la continuité du développement embryonnaire hors de l’œuf. La première mue permet à la larve de devenir autonome tant en termes d’alimentation que de déplacement.
Chez les insectes, le stade pronymphe est présent dans des ordres « primitifs » tel que celui des lépismes (amétabole) (jour 3 et 4 après l’émergence de l’œuf), et très peu observable chez les hémimétaboles. Ainsi, chez les Odonates, les Ephéméroptères (éphémères), les Plécoptères (perles) et les Orthoptères, le stade pronymphal s’étend de la fin du développement embryonnaire à quelques minutes, voire quelques heures, après l’émergence de l’œuf. Pour les autres, comme les Hémiptères, le stade pronymphal correspond uniquement à la période de l’émergence de l’œuf (figure 2 ci-dessous).
La larve pronymphe présente des proportions morphologiques singulières reflétant probablement un compromis entre taille des appendices et confinement lié au faible espace disponible dans l’œuf. Pour optimiser le volume disponible dans l’œuf, la cuticule (exosquelette chez les insectes) de la pronymphe est flexible, les mandibules non sclérifiées et les ailes présentes sous forme de bourgeons.
Contrôle du développement post-embryonnaire
Les américains James W. Truman et Lynn M. Riddiford (1999) (Université de Washington – Seattle) ont mené des études endocrinologiques embryonnaires et post-embryonnaires sur plusieurs espèces d’insectes, leurs résultats appuient l’hypothèse de Berlese : la larve des holométaboles correspondrait au stade pronymphe des hémimétaboles.
Les processus de la mue et de la métamorphose sont gouvernés par 2 familles d’hormones : les ecdystéroïdes comme l’Ecdysone (appelée « Hormone de mue ») et les Hormones Juvéniles (JH).
De nombreux processus physiologiques sont régulés par la JH. Elle a d’abord un rôle de maintien du statu quo ; c’est-à-dire que, de par sa présence dans l’hémolymphe au cours de la mue, elle maintient les caractères larvaires de l’insecte. L’insecte s’extirpe de son ancienne cuticule et gagne en volume par l’extension de la nouvelle encore flexible.
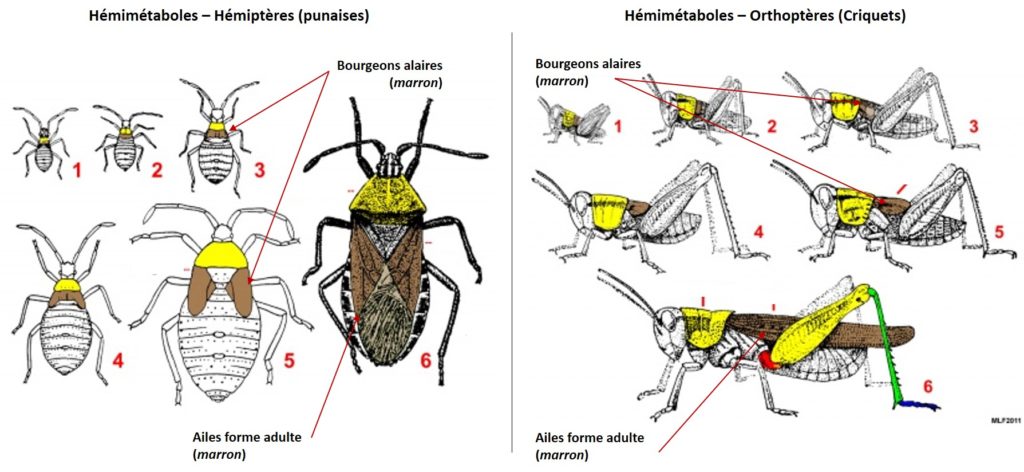
L’Ecdysone, qui est responsable du phénomène de mue, répète l’état de développement précédent et une mue larvaire est réalisée. En revanche, si la JH est absente, l’Ecdysone permet la métamorphose, ou mue imaginale, aboutissant à un individu adulte avec apparition des organes reproducteurs et des ailes pour les espèces ailées (figures 3 et 4).
La JH inhibe donc le processus de métamorphose.
Elle joue également un rôle dans la différenciation des castes chez les insectes sociaux (ouvriers et soldats par exemple chez les termites). Chez l’adulte, elle intervient dans la régulation de l’ovogenèse et le comportement sexuel.
Chez les Holométaboles, les interactions endocriniennes sont plus complexes. Le processus de la mue est induit par un pic de sécrétion d’ecdystéroïdes (appelé pic d’engagement) et une diminution du taux de JH dans l’hémolymphe. Le pic d’engagement déclenche alors un processus irréversible engageant la larve vers la métamorphose. Un second pic d’ecdystéroïdes apparaît dans les heures suivant le premier, pouvant être 10 fois plus important que le pic d’engagement, (temps variable selon les espèces et les taxons) déclenchant la sécrétion de la cuticule de la pupe (figure 4).
La synthèse ou non de JH entraîne une réaction en chaîne aboutissant à l’activation et à l’expression de cortèges de gènes différents à l’origine des mécanismes de la mue et de la métamorphose.
Des caractéristiques morphologiques viennent également conforter cette hypothèse : absence de bourgeons alaires chez les pronymphes hémimétaboles et les larves holométaboles, la similitude du développement intermédiaire des pattes entre la larve holométabole et la forme adulte et la pronymphe hémimétabole (figure 3 ci-dessus).
Initiation de la mue et de la métamorphose
Les facteurs responsables de l’initiation de la mue ne sont pas encore bien décrits. La métamorphose est toutefois induite lorsque les ecdystéroïdes sont synthétisées en l’absence de JH.
Ce qui est admis est que la mue est déclenchée lorsque la larve atteint une taille seuil, bien que ce critère soit subjectif.
Par exemple, chez la punaise hématophage Rhodnius, le processus de mue est induit à la suite de la prise d’un très grand volume de sang, où l’extension des organes abdominaux stimule les récepteurs hormonaux ventraux. Chez le sphinx du tabac (Manduca sexta – Sphingidae), la mue est initiée lorsque la chenille atteint une certaine masse. Les mécanismes permettant à la chenille d’évaluer sa masse ne sont pas encore connus, l’étirement de la paroi corporelle se semblerait pas impliqué. Cependant, la métamorphose serait liée à la taille de la tête au début du stade larvaire. Si la largeur de la tête est inférieure à 5mm, la production de JH se poursuit et la larve engage un nouveau stade larvaire. Si la tête dépasse 5mm, la larve (d’une masse supérieure à 5g) s’engage alors dans le stade pupe (= nymphe) à la mue suivante.
Chez d’autres espèces, la masse n’intervient pas dans le processus de mue. Chez certaines espèces de Coléoptères ou de Lépidoptères où les larves s’alimentent de produits stockés comme la farine ou les graines, les larves muent lorsqu’elles sont affamées et deviennent, étrangement, plus petites.
Mue d’une chenille de papillon du genre Heliconius – Panama 2009 – Benoît GILLES
Production des hormones ecdystéroïdes
Les ecdystéroïdes sont synthétisées dans la glande prothoracique suite à la stimulation de l’hormone Prothoracicotrope (PTTH) provenant quant à elle du cerveau par voie nerveuse. Stockée dans une zone spécifique du cerveau (Corpora radiaca), la libération de la PTTH, généralement via le corpus cardiaques (figure 5), résulte du signal neuronal présumé en lien avec la taille de la larve. Chaque période de production d’ecdystéroïdes est précédée par un pic de PTTH. Pendant la phase d’alimentation du dernier stade larvaire chez la chenille du sphinx du tabac, et probablement chez d’autres espèces, la libération de PTTH est inhibée par une forte production de JH dans l’hémolymphe, bien que PTTH soit libérée en présence de JH durant les stades larvaires précédents.
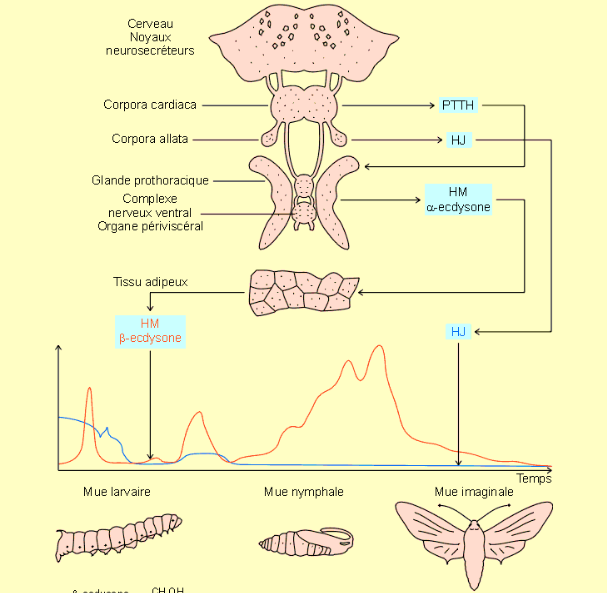
De plus, toujours chez le sphinx du tabac, la libération de PTTH est limitée dans le temps, principalement durant la nuit. La libération de PTTH est alors dite « fermée » du fait que la régulation de l’hormone semble provenir d’un processus interne pouvant être initié à tout moment de la journée mais qui ne peut s’achever qu’à un moment précis de la journée, processus reposant sans doute sur un système d’horloge biologique. Ce mécanisme permet ainsi à la chenille et au papillon de muer durant la nuit lorsque les conditions environnementales sont favorables (température plus faible et humidité plus élevée) et la pression de prédation moindre. Le déclenchement photopériodique de la sécrétion de PTTH est probablement un processus répandu chez les insectes.
Les hormones ecdystéroïdes régulent de nombreuses voies métaboliques en plus de l’activité des cellules épidermiques. Par exemple, le pic d’engagement avant la mue provoque la purge du tube digestif, la régression de tiges dentritiques de certains motoneurones, voire de leur mort. Elle déclenche également la dégénérescence de certains muscles, et la modification de la structuration du système nerveux.
L’action des hormones ecdystéroïdes dépend de la présence ou non de récepteurs spécifiques à la surface de tissus cibles. Les récepteurs peuvent être présents chez un grand nombre de tissus, voire sur l’ensemble des tissus, l’hormone délivrant alors un signal général, ou bien être présents uniquement sur quelques cellules de tissus spécifiques, l’hormone jouant plutôt un rôle de régulateur.
Evolution de la métamorphose
L’apparition et l’évolution de la métamorphose chez les insectes s’explique également par leur écologie. L’avantage que procure la ponte des œufs dans des endroits protégés (sol, écorce) des conditions environnementales et de la prédation, aurait sélectionné ce comportement chez des espèces ancestrales.
Au cours de l’évolution, des adaptations seraient ainsi apparues permettant aux pronymphes holométaboles de se mouvoir sur le site de ponte, mais aussi d’exploiter des ressources alimentaires non disponibles à la nymphe et à l’adulte. Si le milieu offre des conditions favorables, il y a donc un avantage sélectif à ce que la pronymphe s’y développe le plus longtemps possible, et donc à maintenir cette forme durant plusieurs stades larvaires.
La croissance post-embryonnaire (pronymphe) aurait été séparée au départ des stades larvaires et adultes. Le fait que larves et adultes puissent être en compétition pour les mêmes ressources alimentaires aurait privilégié la forme pronymphe : allocation des ressources distinctes entre pronymphe et adulte. La forme nymphe se serait réduite au cours de l’évolution à un seul stade n’ayant plus le besoin de se nourrir, servant uniquement de transition entre les phases larve et adulte : stade de la métamorphose. Il est intéressant de noter qu’un groupe d’hémimétaboles, les trips (Thysanoptères), présente cette phase de transition : 2 stades larvaires suivis par 2 stades « nymphes » ne s’alimentant pas.
Conclusion
L’apparition d’une étape de développement aboutissant à la métamorphose complète est une innovation déterminante dans la diversification des insectes et l’une des clés de leur succès évolutif.
La métamorphose serait apparue à la suite d’un changement temporel de la synthèse de JH et d’une modification du comportement d’oviposition chez la femelle. Au cours de l’évolution, la pronymphe des Hémimétaboles serait devenue la larve des Holométaboles, stade exploitant un habitat totalement différent de celui de la forme adulte où la nymphe serait réduite uniquement à la phase pupe : stade durant lequel la larve ne possède plus la capacité à se nourrir, et où se développent les organes reproducteurs, les ailes et les autres organes caractéristiques de l’adulte.
L’essor des insectes correspond également à l’apparition des premières plantes à fleurs (Angiospermes) durant le Mésozoïque et à leur importante diversification au cours du Crétacé moyen.
Emergence d’un papillon du genre Heliconius – Panama 2009 – Benoît GILLES
Mue imaginale d’un Odonate (Hémimétabole)
Bibliographie
- Truman J. W. & Riddiford L.M. (1999) : The origins of insect metamorphosis. Nature, Vol 401:447-452 l (lien)
- Chapman R.F. (2013) : The Insects – Structure and Function (5ème Edition), 417-435
Recommandation d’ouvrages sur cette thématique
- The Insects : Structure & Function (5th edition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
- Evolution of the Insects (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
- The Insects: An Outline of Entomology (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
- The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
- Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)




Bonjour !
Très bon article de synthèse sur la métamorphose.
Juste une petite précision :
Vous citez blattes, termites et mantes comme appartenant aux isoptères. Cette classification est désormais désuète (elle évolue très vite) et on parle maintenant du super ordre des polyneoptera avec (entre autres) l’ordre des blattodea (blattes et termites) et celui des mantodea (les mantes).
Je vous renvoie au site de l’ITIS pour plus de précision.
Bien cordialement
Sébastien LHOUMEAU