Drosophila suzukii : une petite mouche pas si innocente
Par Gwenaëlle Deconninck & Bréa Simoes-Berton
Drosophila suzukii (Matsumura), aussi connue sous le nom de drosophile à ailes tachetées, est une espèce invasive nuisible originaire d’Asie et désormais présente sur tous les continents. Le mâle est reconnaissable par des tâches noires sur les ailes, qui lui ont donné son surnom (figure 1). Sa capacité d’invasion, son cycle de vie rapide, ses adaptations multiples pour survivre aux nouvelles conditions auxquelles elle fait face, mais aussi les dommages importants qu’elle cause aux cultures, font d’elle un modèle d’actualité pour les études fondamentales et appliquées.

Biologie et écologie
Contrairement à d’autres espèces de drosophiles, D. suzukii pond ses œufs dans des fruits mûrs (figure 2). Pour ce faire, elle utilise un ovipositeur dentelé (organe abdominal permettant la ponte sur ou à l’intérieur d’un fruit, d’un hôte ou du sol par exemple – également appelé oviscape) qui lui permet de percer la peau des fruits en cours de maturation et d’y déposer ses œufs. La femelle pond généralement un à trois œufs par fruit et peut pondre jusqu’à plus de 350 œufs au cours de sa vie.
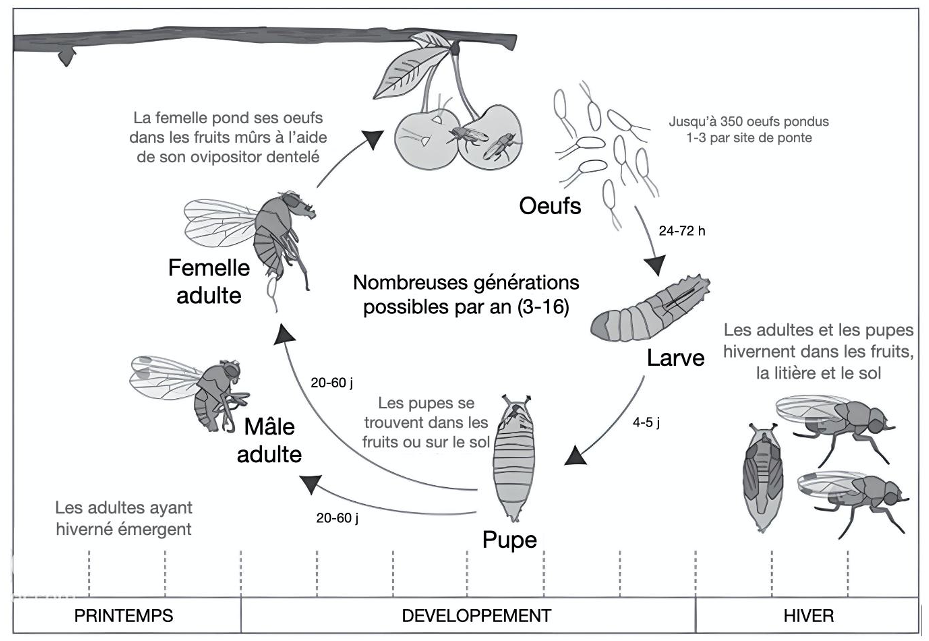
Drosophila suzukii est une espèce particulièrement polyphage (Poyet et al, 2015). Elle est capable de se développer dans une multitude de fruits, aussi bien cultivés que sauvages. Dans des milieux dits hétérogènes, la polyphagie peut être une stratégie adaptative. En l’occurrence, elle permet à D. suzukii de trouver des ressources tout au long de l’année. En hiver, elle va pouvoir utiliser des fruits comme le gui pour se réfugier et hiverner sous forme de pupe, et ainsi avoir de premiers adultes émergeant à l’arrivée du printemps.
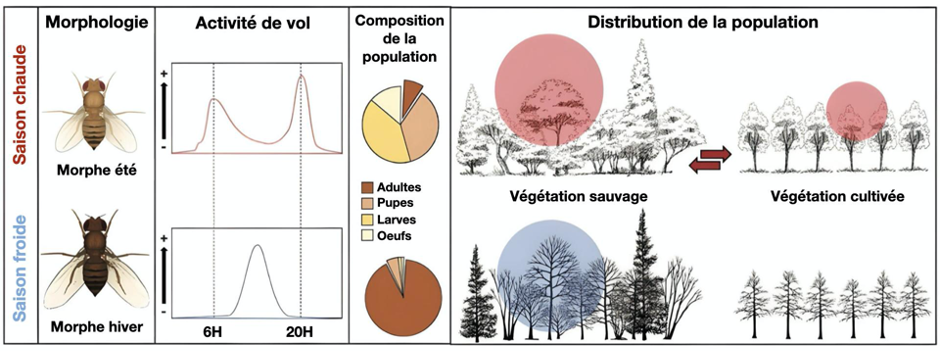
La polyphagie n’est pas la seule stratégie adaptative développée par D. suzukii face aux changements saisonniers. Au cours de l’année, son morphotype, son activité de vol, la composition de sa population vont changer (figure 3). Durant la saison chaude, son morphe « été » peu mélanisé lui permettra de moins subir le réchauffement dû aux rayonnements solaires. Une activité de vol matinale et en soirée lui évitera les températures les plus chaudes de la journée. Ses sites de ponte seront partagés entre les fruits cultivés et les fruits sauvages.
A l’inverse, durant la période hivernale, le morphe « hiver » mélanisé lui permettra de capter les rayons solaires pour se réchauffer et son activité de vol se fera aux moments les plus chauds de la journée. Sa population sera majoritairement composée d’adultes qui se protégeront du froid en trouvant des refuges microclimatiques dans la végétation sauvage.
Son cycle de vie rapide, sa polyphagie et ses multiples adaptations à la saisonnalité font de D. suzukii une espèce particulièrement à même de causer des dégâts importants aussi bien sur la biodiversité que sur les cultures fruitières. D’autant plus qu’il ne s’agit pas d’une espèce locale, mais invasive en Europe.
Une espèce invasive et nuisible
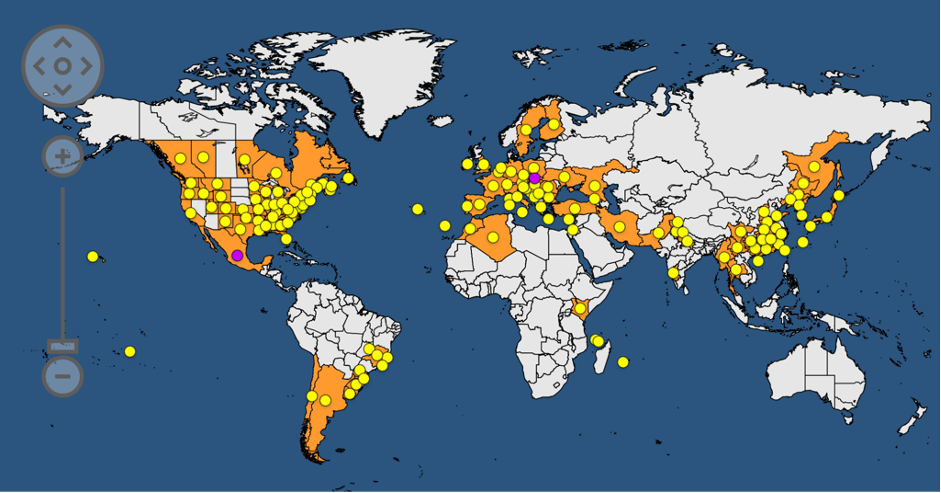
Drosophila suzukii est originaire d’Asie du sud-est. Depuis les années 2000, son territoire n’a cessé de s’étendre (figure 4). Elle a été détectée pour la première fois en Europe et en Amérique du nord en 2008, en Amérique du Sud en 2013, et plus récemment en Afrique du nord en 2017 (Tait et al, 2021). Son expansion a été favorisée par la mondialisation avec l’augmentation des transports de marchandises et humains, mais aussi par les changements globaux. Cette espèce est capable de survivre au transport quel que soit son stade de vie, aussi bien à l’intérieur des fruits qu’au sein des containers.
Sa présence sur les territoires engendrent d’importantes pertes économiques pour les fruiticulteurs. En effet, comme mentionné plus haut, D. suzukii pond ses œufs dans des fruits en mûrissement ou mûrs. La lésion du fruit permet l’entrée de bactéries et champignons qui vont accélérer le pourrissement et le rendre impropre à la consommation ou à la vente. Chaque année, ce sont des pertes allant jusqu’à 80 % de la production avec des coûts économiques allant jusqu’à 500 millions de dollars aux États-Unis par exemple (Young et al, 2018).
Extrêmement polyphage, elle va profiter à chaque saison des nouveaux fruits cultivés à sa disposition, aussi bien des baies que des fruits à noyaux : cerises, fraises, framboises, mûres, myrtilles, kiwis, nectarine, raisin, etc, pour n’en citer que quelques-uns (Poyet et al, 2015 ; Cai et al, 2019). Sur certains fruits, agissant comme pionnière, elle va engendrer l’infestation et la prolifération d’autres espèces, comme Drosophila melanogaster dans les vignobles (Rombaut et al, 2017).
Au-delà des dégâts causés aux producteurs, D. suzukii peut perturber la structure et le fonctionnement des écosystèmes. Même si peu de littérature est disponible à ce sujet, l’arrivée de D. suzukii engendre de fait une modification des flux d’énergie au sein des chaînes trophiques (Poyet et al, 2015) : les fruits qu’elle va utiliser pour le développement de sa descendance ne pourront pas servir à la reproduction de la plante, ne pourront pas servir de site de développement pour d’autres espèces, et ne pourront pas non plus servir de ressource trophique pour des micro-mammifères ou des oiseaux.
Pour lutter contre ce ravageur, des moyens de lutte variés sont développés, notamment dans le cadre de lutte intégrée.
Moyens de lutte
Les larves de D. suzukii peuvent être retrouvées à tous les stades de maturation des fruits, depuis le fruit en cours de maturation, jusqu’au fruit en cours de pourrissement, en passant par le fruit mûr (Bal et al, 2017). Les composts et les fruits tombés au sol sont aussi une source de prolifération des individus. Il est donc très difficile de lutter contre l’infestation des fruits par D. suzukii et des méthodes intégrées sont nécessaires pour limiter au maximum les dégâts occasionnés.
Elles se divisent en quatre catégories principales : la modification des pratiques culturales, le contrôle biologique, le contrôle chimique et la manipulation du comportement (Tait et al, 2021, figure 5).
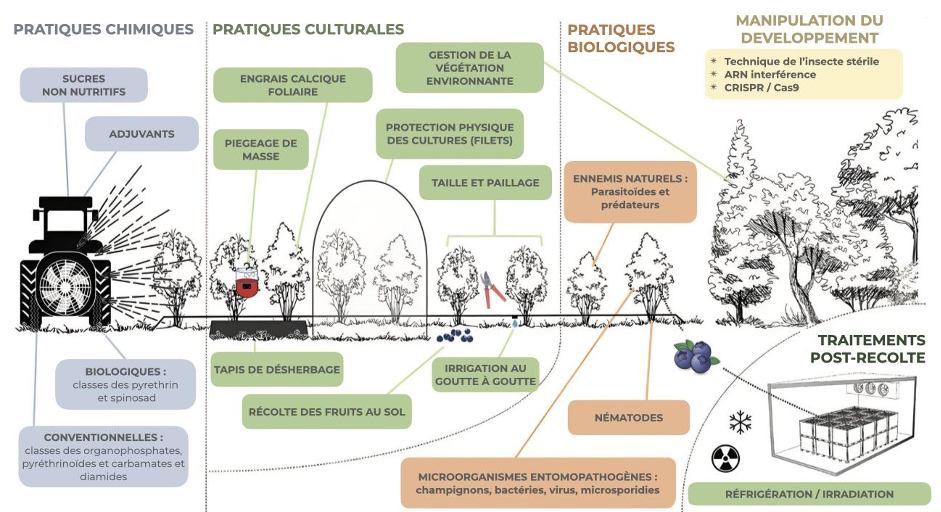
La modification des pratiques culturales consiste à utiliser des moyens physiques pour prévenir l’infestation des fruits par D. suzukii. Parmi les moyens utilisés, peuvent être cités l’utilisation de filets à maille fine (< 0,98 mm) pour couvrir les cultures, l’élimination de plantes sauvages alentours dans lesquelles D. suzukii peut se réfugier, la modification des méthodes d’irrigation pour rendre les conditions moins favorables au développement de D. suzukii, etc. Les méthodes physiques, bien que peu impactantes pour l’environnement, demandent généralement l’acquisition de matériels coûteux et un temps supplémentaire aux fruiticulteurs qui peuvent les rendre moins susceptibles d’être utilisées.
Le contrôle biologique consiste à utiliser des prédateurs, des parasitoïdes ou encore des entomopathogènes pour freiner la prolifération du ravageur. Cette approche peut se faire soit en protégeant les ennemis naturels locaux, en augmentant le nombre d’ennemis naturels présents, ou encore en introduisant de nouveaux ennemis naturels provenant de l’aire de distribution native du ravageur. Pour D. suzukii, les prédateurs (araignées, forficules, coléoptères, etc) n’ont que peu d’impact sur les populations puisque les stades oeuf, larve et pupe sont cachés dans les fruits. Les parasitoïdes généralistes locaux sont également peu efficaces, puisque D. suzukii y est très résistante. Dans son aire native, des parasitoïdes spécialistes sont efficaces (e.g., Ganapsis brasiliensis et Leptopilina japonica) et pourraient être envisagés comme prédateurs introduits. Enfin, l’utilisation d’entomopathogènes (i.e., bactéries, virus, champignons) est prometteuse mais nécessite de poursuivre les recherches.
A l’heure actuelle, la plupart des fruiticulteurs s’appuient sur l’utilisation d’insecticides pour protéger leurs fruits. Les épandages respectent des calendriers précis en fonction de la durée de protection conférée par les molécules et des conditions météorologiques. Les plus utilisées sont des classes des organophosphatés, pyréthrinoïdes, carbamates et diamides. Le spinosad est aussi utilisé en agriculture biologique. Les épandages doivent être répétés régulièrement puisque la pluie va diminuer leur efficacité. Au cours de la saison de production, les classes de molécules utilisées sont alternées pour limiter la résistance aux insecticides. Les insecticides sont les plus utilisés actuellement mais entraînent des coûts économiques et écologiques non négligeables.
La manipulation du comportement peut se faire au travers de différentes méthodes. L’utilisation de pièges attractifs peut permettre de limiter la densité d’adultes dans les champs, mais n’est pas forcément adaptée à de grandes cultures. A l’inverse, des odeurs repoussantes pour les adultes peuvent être utilisées. Par exemple, la mise en place de plantes comme l’alysson maritime (Lobularia maritima) permet de réduire l’infestation de framboises en laboratoire et dans des expériences en cages.
Une dernière méthode pour laquelle les recherches sont en plein essor pour lutter contre D. suzukii est celle de la technique de l’insecte stérile. Utilisée pour de nombreux autres insectes ravageurs (e.g., mouche tsé-tsé, lucilie bouchère, etc), elle a montré son efficacité par le passé. La technique consiste à produire en masse des individus de l’espèce ciblée, de les stériliser par radiations ionisantes et de les relâcher à intervalles réguliers dans les environnements infestés – les individus stérilisés vont ainsi s’accoupler avec les individus stériles sauvages sans produire de descendance. L’utilisation de la bactérie Wolbachia (lien), connue pour manipuler la reproduction chez certains insectes, est aussi prometteuse. Les recherches sont toujours en cours afin de pouvoir déployer cette méthode pour lutter contre D. suzukii.
De nombreuses méthodes de lutte existent mais elles ne sont pas destinées à être utilisées seules, bien au contraire. En combinant ces différentes méthodes dans un objectif de lutte intégrée, l’efficacité contre le ravageur est potentiellement démultipliée.
Intérêt scientifique
Les paragraphes ci-dessus soulèvent déjà certaines questions autour de la compréhension de l’invasion de D. suzukii, de sa niche écologique, de l’amélioration des moyens de lutte contre ce ravageur, et bien d’autres. Les recherches se multiplient sur ce modèle particulièrement intéressant qui permet de lever des fonds pour limiter les dégâts aux cultures et les pertes financières associées.
Nous avons choisi de vous présenter les recherches que nous menons dans le cadre du projet ANR DroThermal « Qu’est-ce qui fait de Drosophila suzukii un envahisseur si efficace ? Une analyse intégrative de son écologie thermique ». Quatre laboratoires français se sont associés pour répondre à cette question :
- Le laboratoire Écosystème, Biodiversité, Évolution (Écobio, UMR6553) à Rennes
- Le laboratoire Écologie et Dynamique des Systèmes Anthropisés (EDYSAN, UMR7058) à Amiens
- L’Institut de Recherche sur la Biologie de l’Insecte (IRBI, UMR7261) à Tours
- Le Laboratoire de Biologie et Biométrie Évolutive (LBBE, UMR5558) à Lyon
Chaque laboratoire travaille sur des aspects complémentaires qui permettent d’améliorer les connaissances fondamentales sur D. suzukii mais aussi qui visent à développer des modèles prédictifs innovants de dynamique des populations, qui seront utiles aux programmes de gestion durable contre ce ravageur.
- Ecobio – Variations thermiques
Dans la nature, D. suzukii va faire face à des environnements thermiques fluctuants, aussi bien à l’échelle des jours que des saisons. Les températures varient en fonction des heures de la journée, de la période de l’année, et ces variations ne sont pas toujours linéaires puisque des événements extrêmes peuvent se produire. Parfois, ces fluctuations vont permettre aux individus de survivre à des températures auxquelles elles n’auraient pas survécu s’ils avaient été exposés pendant une durée plus longue (Petavy et al, 2001). Cette sous-tâche du projet vise à déterminer les conséquences phénotypiques et physiologiques de ces variations de températures pour mieux comprendre et prédire la tolérance à la température de D. suzukii. Un intérêt particulier est porté sur la période hivernale, pour étudier les adaptations mises en place par D. suzukii pour survivre aux conditions froides extrêmes.
Dans la nature, la température environnante fluctue. La durée et l’intensité de ces fluctuations dépendent de l’échelle temporelle étudiée et influencent la mise en place de mécanismes écophysiologiques chez D. suzukii.
Au laboratoire, pour étudier précisément cet effet, les chercheurs contrôlent les paramètres environnementaux (taux d’humidité, photopériode, taux de lumière, etc) en utilisant des chambres climatiques (ou “incubateurs”, figure 6). Celles-ci permettent de tester l’effet d’une température, souvent constante, et fournissent des conditions “idéales”/extrêmement contrôlées, très éloignées des conditions naturelles. Depuis quelques années, certains chercheurs intègrent de la variation dans leurs protocoles en choisissant d’implémenter des fluctuations de températures programmées dans les incubateurs. Cette question du choix de niveau de complexité à adopter dans l’élaboration du design expérimental fait débat au sein de la communauté scientifique de la biologie thermique, certains attestant que les designs les plus simples sont les plus représentatifs (von Schmalensee et al, 2021), d’autres incitant à se tourner vers des designs plus élaborés (Kingsolver et al, 2015).

Cet axe du projet s’intéresse à cette problématique et tend à étudier l’effet de la variation thermique selon différentes échelles temporelles chez D. suzukii en comparant trois niveaux de complexité expérimentale : un design expérimental simple (conditions constantes en incubateur), un design expérimental élaboré (conditions fluctuantes en incubateur) et un design expérimental complexe (conditions naturelles sur le terrain). Les expériences ont été menées en laboratoire en incubateurs et au sein du jardin expérimental rattaché au laboratoire Ecobio. De grandes cages en bois ont été construites et grillagées, pour empêcher d’autres insectes de s’infiltrer, et une toile isolante a été installée sur les parties ajourées des cages pour empêcher un effet de serre dû aux rayonnements du soleil (figure 6).
Les premiers résultats montrent que, dans certains cas, les designs les plus simples peuvent être pertinents. Les résultats obtenus suite à une étude de la tolérance au froid chez des adultes D. suzukii, ont montré qu’une exposition constante à une température basse induit les mêmes réponses de tolérance qu’une exposition en milieu naturel pendant la saison automnale. Toutefois, d’autres résultats montrent que les designs les plus simples ne sont pas suffisants dans d’autres cas et la mise en place de designs élaborés est nécessaire pour obtenir des résultats pertinents écologiquement. Par exemple, une étude de l’effet de la température sur le temps de développement de D. suzukii montre que pour certaines conditions extrêmes chaudes et froides, une exposition à des conditions fluctuantes est primordiale pour obtenir des résultats fiables.
- EDYSAN – Variations trophiques
Comme indiqué plus haut, D. suzukii est une espèce extrêmement polyphage : elle est capable d’assurer son développement dans une multitude de fruits. En hiver, elle doit malgré tout faire face à une diminution drastique des fruits disponibles (Hamby et al, 2016). Ils sont moins nombreux, et potentiellement de moins bonne qualité nutritionnelle. Or, la qualité de l’alimentation va impacter la tolérance au froid de l’espèce. Cette sous-tâche vise donc à identifier les ressources disponibles pour D. suzukii en hiver, et à déterminer leur contribution à sa survie en fonction de différentes conditions de température.
Une première étude préliminaire recensait la disponibilité et l’utilisation des fruits tout au long de l’année par D. suzukii (Poyet et al, 2015). En hiver, les espèces dans lesquelles elle pond sont moins nombreuses, et la proportion de celles ne permettant pas son développement complet augmente (figure 7).
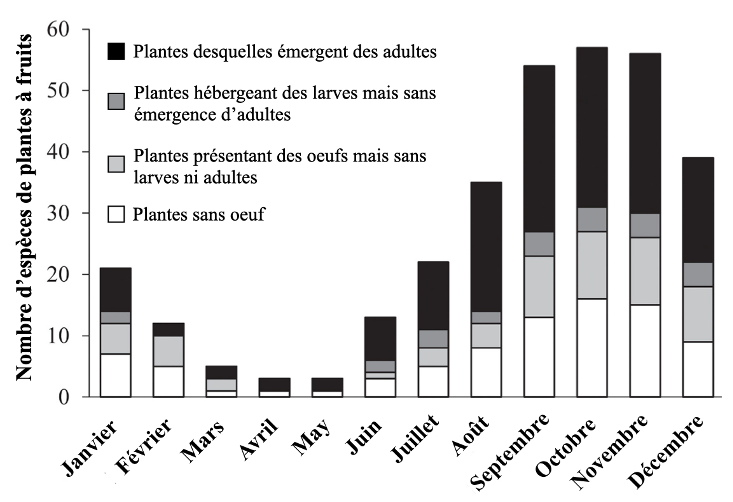
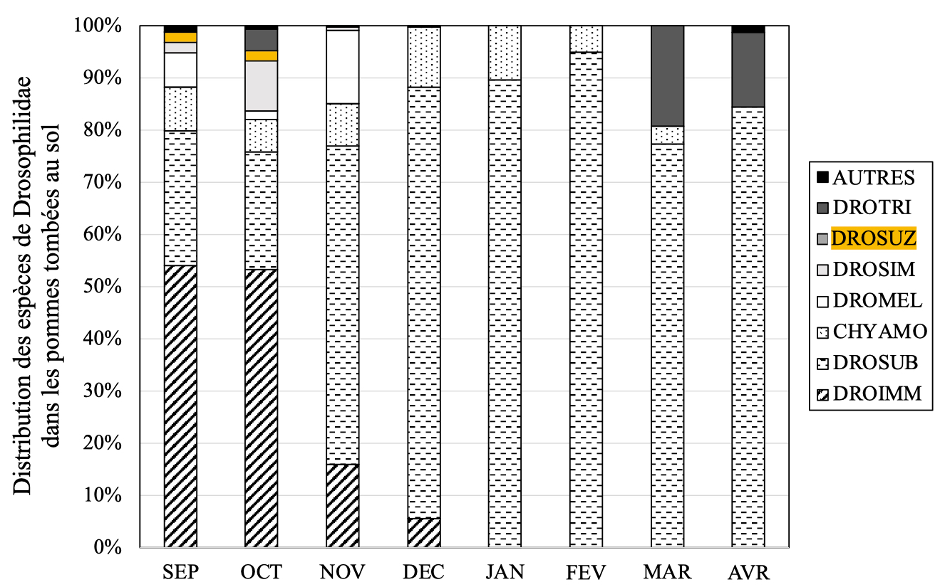
Jusqu’à présent négligés, les fruits tombés au sol peuvent également constituer un réservoir pour D. suzukii. C’est ce que nous avons montré au travers de l’étude de la communauté des Drosophilidae retrouvés dans les pommes tombées au sol, qui se décomposent tout au long de l’hiver (Boulembert, Deconninck, et al, en préparation). Celles-ci permettent à de nombreuses espèces de se réfugier et de survivre aux saisons plus froides (automne et hiver), dont D. suzukii durant les premiers mois (figure 8).
La suite de cette sous-tâche s’intéressera à d’autres fruits d’hiver utilisés par D. suzukii comme la plante ornementale Aucuba japonica, le gui Viscum album ou encore l’arbuste Elaeagnus x ebbengei. En particulier, l’impact de la qualité nutritionnelle de ces fruits sur le développement et la performance de D. suzukii sera étudié.
- IRBI – Variations et thermorégulation
Dans des environnements dits hétérogènes, comprenant une multitude de microhabitats avec des conditions de température et d’humidité différentes, la capacité de thermorégulation comportementale est essentielle. Elle consiste à se déplacer d’un environnement défavorable vers un environnement favorable pour assurer sa survie ou améliorer ses performances. Pour l’organisme, elle implique d’avoir des microclimats à sa disposition, de pouvoir exploiter ces zones, et qu’elles aient un effet à court et long terme sur des traits physiologiques (Woods et al, 2015). Cette sous-tâche vise à caractériser la capacité de D. suzukii à utiliser la thermorégulation comportementale pour survivre et améliorer ses performances.
Pour étudier la thermorégulation comportementale, le trait mesuré est la préférence thermique. Il s’agit de la température qu’un individu va choisir, par exemple pour optimiser ses performances ou à l’inverse pour économiser ses réserves énergétiques. Pour mesurer cette préférence thermique, différents dispositifs existent. A l’IRBI, un gradient thermique à une dimension a été choisi (figure 9).
Il s’agit d’une plaque d’aluminium dans laquelle sont creusées des “rampes” où les mouches peuvent se déplacer librement dans une direction. On s’intéresse à la variation de cette préférence thermique en fonction de différents paramètres, comme le stade de développement (larves ou adultes), la température de développement, l’âge des individus ou leur sexe. Les premiers résultats montrent que cette préférence thermique est extrêmement variable (et donc plastique). Tous les paramètres étudiés ont engendré des modifications.
Particulièrement intéressants, ces résultats pourraient expliquer en partie pourquoi D. suzukii est une espèce invasive si performante.
En effet, si elle est capable de moduler à ce point sa préférence thermique, et donc ses capacités de thermorégulation comportementale, elle peut s’adapter à la plupart des environnements qu’elle va rencontrer.

- LBBE – Interactions microbiennes symbiotiques
Les interactions symbiotiques avec des bactéries se retrouvent dans tout le règne animal. Chez les insectes, ces symbioses peuvent être aussi bien facultatives qu’obligatoires, et aussi bien bénéfiques que délétères. Par exemple, certaines bactéries vont complémenter leur hôte en vitamines, alors que d’autres vont manipuler leur reproduction. Certaines bactéries encore vont avoir des rôles différents en fonction des hôtes qu’elles infectent. C’est le cas de Wolbachia, bactérie présente chez plus de 50 % des espèces d’insectes (Weinert et al, 2015). Drosophila suzukii est infectée par Wolbachia, mais son action reste indéterminée. L’objectif de cette sous-tâche consiste à décrire la communauté symbiotique présente chez D. suzukii et à déterminer si elle joue un rôle dans sa réponse aux stress thermiques.
Pour cette sous-tâche nous avons exploré certains effets potentiels que pourrait avoir Wolbachia chez D. suzukii, et en particulier les effets sur la survie, la fécondité, le temps de développement, la résistance face à un changement de milieu nutritionnel, le métabolisme, les réserves énergétiques, ou encore la thermorégulation comportementale. Jusqu’à présent, les résultats obtenus pour ces différents traits ne laissent pas présumer d’effet très net. Wolbachia semble être plutôt bénéfique pour D. suzukii puisqu’elle augmente sa survie dans le cadre d’un stress nutritionnel (changement de milieu de développement).
Les autres effets ne vont s’observer que dans des conditions bien particulières. Par exemple, les individus infectés par Wolbachia vont voir leur métabolisme diminuer en condition de températures fluctuantes, mais pas à température constante. De la même manière, les individus infectés par Wolbachia vont avoir des réserves en glycogène plus importantes que les individus qui ne le sont pas, mais uniquement en régime fluctuant ou à température élevée. Les recherches se poursuivent pour mettre en lien tous ces résultats et éclaircir le rôle de cette bactérie sur son hôte.
Conclusion
Les recherches autour de D. suzukii s’intensifient, aussi bien du point de vue fondamental qu’appliqué.
Dans le monde entier, les chercheurs acquièrent de nouvelles connaissances sur ce modèle et la France occupe un rôle central dans les avancées scientifiques.
Seules les approches dites intégratives, c’est à dire qui prennent en compte différents niveaux d’informations (e.g., biologique, physiologique, biochimique) et leurs interactions au sein de l’organisme et de l’écosystème, seront à même de mieux saisir les raisons du succès invasif de D. suzukii et ses enjeux aussi bien agronomiques que pour les écosystèmes naturels. Les collaborations entre chercheurs sont essentielles pour y parvenir.
AuteursGwenaëlle Deconninck
Bréa Simoes-Berton
|
 Ma passion pour la recherche est née sur une île grecque, durant un stage où j’inventoriais les espèces d’orchidées méditerranéennes présentes, tout en cherchant à améliorer les conditions de culture in vitro pour la préservation de ces fleurs menacées. Par la suite, j’ai étudié la spécificité de la pollinisation d’espèces de Ceropegia en sympatrie par des diptères. Pour finalement, en thèse, m’intéresser à la préférence thermique et trophique d’une espèce invasive, D. suzukii. Je me passionne pour la compréhension des mécanismes qui régissent les phénomènes écologiques au sein des écosystèmes, notamment terrestres. Les interactions plantes-insectes me fascinent par leur diversité et leur complexité, tout comme les interactions avec d’autres organismes comme des bactéries ou des champignons.
Ma passion pour la recherche est née sur une île grecque, durant un stage où j’inventoriais les espèces d’orchidées méditerranéennes présentes, tout en cherchant à améliorer les conditions de culture in vitro pour la préservation de ces fleurs menacées. Par la suite, j’ai étudié la spécificité de la pollinisation d’espèces de Ceropegia en sympatrie par des diptères. Pour finalement, en thèse, m’intéresser à la préférence thermique et trophique d’une espèce invasive, D. suzukii. Je me passionne pour la compréhension des mécanismes qui régissent les phénomènes écologiques au sein des écosystèmes, notamment terrestres. Les interactions plantes-insectes me fascinent par leur diversité et leur complexité, tout comme les interactions avec d’autres organismes comme des bactéries ou des champignons. Passionnée depuis toujours d’animaux et de nature, je me suis souvent posé beaucoup de questions sur la manière dont fonctionne l’environnement qui nous entoure. C’est tout naturellement que j’ai suivi une licence puis un master d’écologie et de biologie. Les trois stages que j’ai faits m’ont confortée dans l’idée que j’avais trouvé ma voie, et c’est le stage de master 2 qui a été une véritable vocation. Ce stage était une étude de l’effet des îlots de chaleur urbaine sur l’écologie thermique et la thermopréférence du gendarme Pyrrhocoris apterus. En effet, j’affectionne particulièrement l’étude de l’effet de la température sur la faune en lien avec des problématiques centrées autour des effets du changement climatique. Aujourd’hui, j’ai la chance de faire une thèse dont les travaux de recherche se concentrent sur l’effet des variations temporelles de la température sur les traits phénotypiques et écophysiologique chez D. suzukii Ce sujet de thèse est d’autant plus fascinant que la drosophile à ailes tachetées est un très petit insecte qui a pourtant réussi à coloniser différents habitats dans le monde, régis par différents climats. Pour la suite de ma carrière, je serai très intéressée de continuer à étudier cet organisme au cours d’un contrat post-doctoral par exemple.
Passionnée depuis toujours d’animaux et de nature, je me suis souvent posé beaucoup de questions sur la manière dont fonctionne l’environnement qui nous entoure. C’est tout naturellement que j’ai suivi une licence puis un master d’écologie et de biologie. Les trois stages que j’ai faits m’ont confortée dans l’idée que j’avais trouvé ma voie, et c’est le stage de master 2 qui a été une véritable vocation. Ce stage était une étude de l’effet des îlots de chaleur urbaine sur l’écologie thermique et la thermopréférence du gendarme Pyrrhocoris apterus. En effet, j’affectionne particulièrement l’étude de l’effet de la température sur la faune en lien avec des problématiques centrées autour des effets du changement climatique. Aujourd’hui, j’ai la chance de faire une thèse dont les travaux de recherche se concentrent sur l’effet des variations temporelles de la température sur les traits phénotypiques et écophysiologique chez D. suzukii Ce sujet de thèse est d’autant plus fascinant que la drosophile à ailes tachetées est un très petit insecte qui a pourtant réussi à coloniser différents habitats dans le monde, régis par différents climats. Pour la suite de ma carrière, je serai très intéressée de continuer à étudier cet organisme au cours d’un contrat post-doctoral par exemple.
Bibliographie
- Bal, H. K., Adams, C., & Grieshop, M. (2017) : Evaluation of Off-season Potential Breeding Sources for Spotted Wing Drosophila (Drosophila suzukii Matsumura) in Michigan. Journal of Economic Entomology, 110(6), 2466–2470 (lien) https://doi.org/10.1093/jee/tox252
- Boulembert, M., Deconninck, G., Eslin, P., Couty, A., Dubois, F., Gallet‐Moron, E., Pincebourde, S., & Chabrerie, O. (En préparation) : Local and landscape factors shape Drosophilidae diversity in decaying apple fruits across seasons
- Cai, P., Song, Y., Yi, C., Zhang, Q., Xia, H., Lin, J., Zhang, H., Yang, J., Ji, Q., & Chen, J. (2019) : Potential host fruits for Drosophila suzukii: olfactory and oviposition preferences and suitability for development. Entomologia Experimentalis et Applicata, 167(10), 880–890 (lien) https://doi.org/10.1111/eea.12840
- Hamby, K. A., E. Bellamy, D., Chiu, J. C., Lee, J. C., Walton, V. M., Wiman, N. G., York, R. M., & Biondi, A. (2016) : Biotic and abiotic factors impacting development, behavior, phenology, and reproductive biology of Drosophila suzukii. Journal of Pest Science, 89(3), 605–619 (lien) https://doi.org/10.1007/s10340-016-0756-5
- Kingsolver, J., Higgins, J.K., Augustine, K.E. (2015) : Fluctuating temperatures and ectotherm growth: distinguishing non-linear and time-dependent effects. Journal of experimental biology, 218(14) (lien) https://doi.org/10.1242/jeb.120733
- Petavy, G., David, J. R., Gibert, P., & Moreteau, B. (2001) : Viability and rate of development at different temperatures in Drosophila: a comparison of constant and alternating thermal regimes. Journal of Thermal Biology, 26(1), 29–39 (lien) https://doi.org/10.1016/S0306-4565(00)00022-X
- Poyet, M., Le Roux, V., Gibert, P., Meirland, A., Prévost, G., Eslin, P., & Chabrerie, O. (2015) : The wide potential trophic niche of the asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE, 10(11) (lien) https://doi.org/10.1371/journal.pone.0142785
- Rombaut, A., Guilhot, R., Xuéreb, A., Benoit, L., Chapuis, M. P., Gibert, P., & Fellous, S. (2017) : Invasive Drosophila suzukii facilitates Drosophila melanogaster infestation and sour rot outbreaks in the vineyards. Royal Society Open Science, 4(3) (lien) https://doi.org/10.1098/rsos.170117
- von Schmalensee, L., Gunnarsdottir, K.H., Naslund, J., Gotthard, K., Lekmann, P. (2021) : Thermal performance under constant temperatures can accurately predict insect development times across naturally variable microclimates. Ecology Letters, 24(8) (lien) https://doi.org/10.1111/ele.13779
- Tait, G., Mermer, S., Stockton, D., Lee, J., Avosani, S., Abrieux, A., Anfora, G., Beers, E., Biondi, A., Burrack, H., Cha, D., Chiu, J. C., Choi, M.-Y., Cloonan, K., Crava, C. M., Daane, K. M., Dalton, D. T., Diepenbrock, L., Fanning, P., … Walton, V. M. (2021) : Drosophila suzukii (Diptera: Drosophilidae): A Decade of Research Towards a Sustainable Integrated Pest Management Program . Journal of Economic Entomology, 114(5), 1950–1974 (lien) https://doi.org/10.1093/jee/toab158
- Weinert, L. A., Araujo-Jnr, E. V., Ahmed, M. Z., & Welch, J. J. (2015) : The incidence of bacterial endosymbionts in terrestrial arthropods. Proceedings of the Royal Society B: Biological Sciences, 282(1807), 20150249 (lien) https://doi.org/10.1098/rspb.2015.0249
- Woods, H. A., Dillon, M. E., & Pincebourde, S. (2015) : The roles of microclimatic diversity and of behavior in mediating the responses of ectotherms to climate change. Journal of Thermal Biology, 54, 86–97 (lien) https://doi.org/10.1016/j.jtherbio.2014.10.002
- Young, Y., Buckiewicz, N., & Long, T. A. F. (2018) : Nutritional geometry and fitness consequences in Drosophila suzukii, the Spotted-Wing Drosophila. Ecology and Evolution, 8(5), 2842–2851 (lien) https://doi.org/10.1002/ece3.3849




