Des gènes à l’origine de traits nouveaux donnant accès à un nouvel environnement
chez la punaise Rhagovelia (Heteroptera, Gerromorpha, Veliidae)
Les innovations évolutives se traduisent par l’apparition de nouvelles structures ou comportements au cours de l’évolution (ex : la fleur, la carapace des tortues, l’aile des insectes (lien), un patron de coloration chez les papillons (lien)). La théorie prédit que ces événements d’apparition sont uniques à chaque innovation et rares. C’est ainsi que les innovations sont partagées par des groupes d’espèces apparentées, des taxons (ex : respectivement, les angiospermes, les oiseaux, les tortues, les insectes, les papillons). L’apparition de nouvelles structures est le premier mécanisme par lequel les innovations participent à la diversification du vivant.
Une innovation est considérée comme clé, importante, lorsqu’elle permet l’adaptation à une nouvelle niche écologique via une nouvelle fonction. La théorie prédit que l’opportunité écologique ainsi créée va permettre la diversification de l’espèce possédant l’innovation en groupe d’espèces. C’est le deuxième mécanisme par lequel les innovations participent à la diversification du vivant. Ainsi, elles façonnent les trajectoires évolutives des lignées dans lesquelles elles apparaissent.
L’importance de certaines de ces innovations pousse les chercheurs à se demander quels sont les mécanismes génétiques et développementaux à leur origine. Ils cherchent à faire le lien entre les changement génétiques, phénotypiques et adaptatifs à un nouveau milieu.
Punaises du genre Rhagovelia à la surface de l’eau (Source : A. Khila, 2017)
Pour tenter de répondre à ces questions, l’équipe de Khila Abderrahman (CNRS-UMR5242, Université de Lyon) a étudié les mécanismes génétiques et les pressions environnementales qui sous-tendent l’évolution de « l’éventail propulseur » (photo 2 ci-dessous), structure exclusive des punaises du genre Rhagovelia (Heteroptera, Gerromorphe, Veliidae) qui leur permet de se propulser à la surface de l’eau.
Leurs résultats mettent en évidence le rôle central de gènes qui sont spécifiques au taxon. L’un d’eux n’est présent que chez les punaises d’eau et l’autre que dans le groupe qui possède l’éventail (le genre Rhagovelia). Ces deux gènes sont impliqués dans le développement de l’éventail. De plus, l’utilisation de tests biomécaniques a prouvé le rôle essentiel de l’éventail dans l’adaptation à un nouvel environnement, agissant ainsi comme une innovation déterminante au plan évolutif.
Biologie et description des Rhagovelia
Le genre Rhagovelia (photo 1 ci-dessous) est constitué de ∼200 espèces dont le cycle biologique se déroule, en général, à la surface de petits courants d’eau rapides – niche écologique pas ou peu accessible à la plupart des autres insectes semi-aquatiques -. Ces punaises aquatiques se déplacent à la surface par des mouvements d’aviron de la paire de pattes médianes où se situent les éventails. Un éventail se compose d’environ 20 branches (ou fanes) primaires chacune, dotée de branches secondaires plus minces (photo 2 ci-contre). Contrairement à Rhagovelia, le genre Stridulivelia, qui lui est étroitement apparenté, ne possède pas d’éventail de propulsion. Bien que les espèces des deux genres occupent les mêmes cours d’eau (espèces sympatriques), les punaises du genre Stridulivelia restent statiques sur les feuilles des plantes et n’effectuant qu’occasionnellement des déplacements rapides (voir vidéo 1).
Photo 1 : Rhagovelia antilleana – les éventails sont visibles sur les pattes médianes (Source : Sciencesnews.org)Photo 2 : éventail propulseur chez Rhagovelia antilleana (Source : Santos et al., 2017) (Modifié par B. gilles)
Le développement embryonnaire de l’éventail débute entre 144 et 210 heures, soit à 35% de la durée totale de l’embryogenèse (à 26°C). Les larves du premier stade éclosent avec des éventails entièrement fonctionnels, renouvelés au cours de chacune de leurs cinq mues, indiquant la persistance du programme de développement du éventail tout au long des stades post-embryonnaires (illustration 1 ci-dessous).
Illustration 1 : Développement de l’éventail au 1er, 3ème et 5ème stade larvaire – augmentation de la taille et du nombre des branches (Source : Santos et al., 2017)
Analyses génétiques
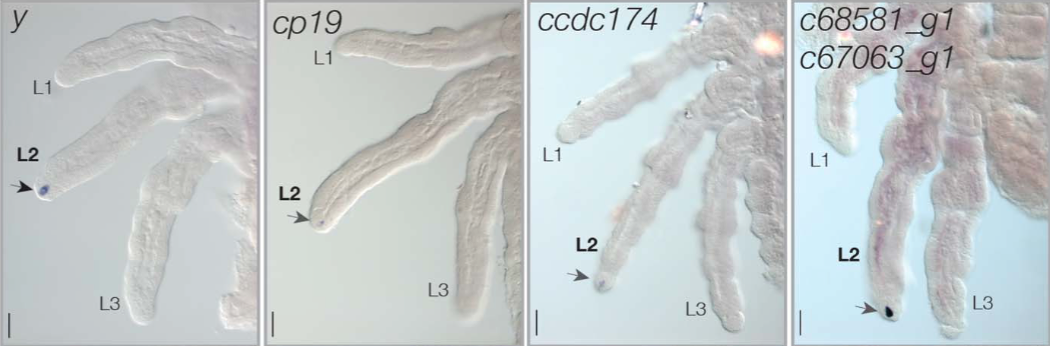
Pour identifier les gènes à l’origine du développement de l’éventail, l’équipe de scientifiques a réalisé un profilage transcriptomique (étude de l’ensemble des ARN messager produits lors du processus de transcription d’un génome) des pattes embryonnaires chez Rhagovelia antilleana. L’étude de l’expression de ces ARNm a permis d’identifier 5 gènes s’exprimant uniquement dans la paire de pattes médianes et à l’endroit où se développe l’éventail : y (yellow), cp19 (cuticular protéine 19), ccdc174 (coiled-coil domain-containing protéine 174) et deux gènes similaires inconnus (c67063_g1 et c68581_g1, respectivement nommé geisha et mother-of-geisha) (illustration 2 ci-dessous). Aucun de ces gènes ne s’exprime dans les pattes antérieures et postérieures de R. antilleana ou de toute autre espèce apparentée comme Stridulivelia tersa et Oiovelia cunucunumana.
Illustration 2 : Expression des 5 gènes à l’apex de la patte médiane de Rhagovelia (Source : Santos et al., 2017)
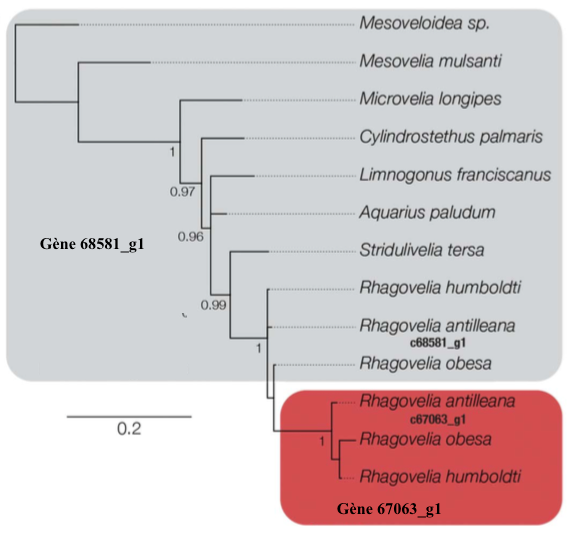
Une recherche d’homologie des séquences liées à geisha et mother-of-geishaa montré que mother-of-geisha possédait un paralogue (gène homologue) retrouvé chez 5 espèces d’Hémiptères (punaises) et une d’Isoptères (termites). Aucun de ces deux gènes ne possède d’homologue dans d’autres Ordres comme les Lépidoptères, les Diptères et les Hyménoptères. Le gène mother-of-geisha possède cependant de proches homologues dans les espèces de punaise d’eau n’ayant pas d’éventail (i.e. autres que Rhagovelia) alors que toutes les espèces de Rhagovelia et seulement celles-ci partagent le gène geisha. Les chercheurs concluent que mother-of-geisha a émergé chez une espèce ancestrale commune à tous les Hémiptères et s’est ensuite dupliqué chez l’ancêtre commun du clade des Rhagovelia pour donner geisha. Dans cet ancêtre commun, la séquence de geisha a évolué très rapidement et a divergé de celle de mother-of-geisha (illustration 3 ci-dessous). C’est ainsi que les gènes ont des séquences très proches mais facilement différentiables dans toutes les espèces de Rhagovelia.
Illustration 3 : reconstruction phylogénétique des deux gènes (Source : Santos et al., 2017) (Modifié par B. Gilles)
L’examen de l’expression de ces deux gènes a révélé que geishaétait exprimé trois fois plus fortement que mother-of-geishadans les pattes médianes. Ainsi, non seulement la séquence protéique de ces gènes a divergé, mais aussi leur niveau d’expression.
Sachant que mother-of-geisha et geisha sont respectivement spécifiques des Hémiptères et des Rhagovelia et qu’ils sont exprimés à l’endroit et au moment du développement de l’éventail, les scientifiques ont souhaité tester s’ils étaient impliqués dans le développement de cette structure. Pour cela, ils ont inhibé l’expression de ces gènes lors du développement des larves. Ainsi, l’inhibition de l’expression des gènes mother-of-geisha et geishaa entrainé de la réduction à la disparition de l’éventail, tout en permettant le développement d’individus viables sans autres modifications phénotypiques d’autres structures (y compris des griffes directement liées à l’éventail) (Illustration 4 ci-contre).
Illustration 4 : Comparaison des phénotype sauvage et suivi d’une inhibition des gènes (Source : Santos et al., 2017) (Modifié par B. gilles)
La suppression du gène y « yellow » a eu pour conséquence le développement du phénotype « éventail jaunâtre » au lieu de couleur brun foncé. Ce gène connu pour intervenir dans la formation de pigments foncés, et conservé au sein du genre Rhagovelia, a donc été recruté pour assombrir la cuticule des éventails.
La surpression des gènes cp19 et ccdc174 connus pour leur implication dans le développement de la cuticule n’a quant à elle pas engendré de perturbation détectable pour des raisons non identifiées.
En résumé, dans cette partie les chercheurs ont établi un lien entre des changements génétiques (apparition de mother-of-geisha, duplication de geisha, évolution rapide de la séquence de geisha)et la génétique du développement de l’éventail (ces deux gènes sont fortement exprimés dans le territoire présomptif de l’éventail, leur expression est nécessaire au développement de l’éventail, yellow a été recruté lors du processus évolutif). Pour finir de reconstruire l’histoire évolutive ils ont tenté de comprendre en quoi ces changements étaient adaptatifs.
Mots de Abderahman Khali
A. Khila sur le terrain (Source : A. Khila)
Vous dirigez l’Unité de recherche « Génomique du développement et évolution », quelles sont les thématiques et les modèles d’études que vous étudiez ?
Je dirige ce laboratoire au sein de l’UMR5242 regroupant 12 autres laboratoires. Dans mon laboratoire, nous nous intéressons aux mécanismes génétiques, évolutifs et écologiques à l’origine de la diversification des espèces. Les modèles d’études sont multiples : le poisson zèbre (Danio rerio), la drosophile (Drosophila melanogaster), la souris domestique, le tribolium (Tribolium castaneum – Coleoptera) et les Gerromorphes (punaises aquatiques).
Quels sont les outils innovants que vos équipes utilisent dans le cadre de leurs recherches ?
Nos équipes utilisent notamment une plateforme de séquençage à haut débit (PSI), le génie génétique et la manipulation des génomes, l’imagerie et microscopie, la modélisation, etc.
Que vont apporter vos recherches fondamentales en entomologie ces prochaines années et décennies ?
L’apport principal est d’accroitre nos connaissances scientifiques. D’autres apports peuvent toucher la sensibilisation du public par rapport aux écosystèmes et leur sauvegarde, et à un degré moindre, la lutte contre les insectes nuisibles.
Etudes du déplacement et du comportement
Un des objectifs majeurs de la biologie évolutive est non seulement d’identifier les mécanismes génétiques sous-jacents aux caractères nouveaux, mais aussi de déterminer l’impact des innovations sur l’histoire évolutive de l’organisme. Afin de déterminer si l’innovation éventail a permis l’adaptation à une nouvelle niche, les chercheurs ont testé si elle pouvait avoir participé à l’invasion des petits cours d’eau où vivent les Rhagovelia. Ainsi, ils se sont amusés à comparer les performances de locomotion par vidéographie à haute vitesse des déplacements à la surface de l’eau d’individus de Rhagovelia : 1) normaux (avec éventail), 2) sans éventail (ablation chirurgicale), 3) avec éventail rudimentaire (inhibition des gènes mother-of-geisha et geisha), 4) Stridulivelia tersa, une espèce proche sans éventail, qui vit aussi sur près de cours d’eau mais s’aventurant rarement à la surface.
Dans un premier temps, les chercheurs ont simplement confirmé que seul l’éventail était affecté par l’inhibition des gènes mother-of-geisha et geisha, et que les rudiments restants comme les griffes se déployaient et se rétractaient normalement.
Les scientifiques ont réalisé des comparaisons des performances locomotrices à la surface d’eau d’individus de ces 4 groupes. La première comparaison a été faite en eau stagnante. Pour cela, Stridulivelia tersa y sont plus rapides que les 3 groupes de Rhagovelia qui sont tous les 3 au même niveau. Cependant, Stridulivelia tersa et les Rhagovelia sans éventail font plus de battements de pattes par seconde. Les chercheurs concluent que sur l’eau stagnante les éventails ne sont pas utilisés pour augmenter la vitesse moyenne mais permettent de diminuer la fréquence des battements.
Ensuite, une deuxième expérience a consisté à obliger les groupes d’individus à ramer contre un courant d’eau lent (0,2m/s) et rapide (0,3m/s), simulant l’environnement des R. antilleana. Bien que les individus de S. tersa aient été plus rapides en eau stagnante, ils ont été emportés par les courant lents et rapides. Les individus R. antilleana normaux (témoins) ont quant à eux remonté les deux types de courant sans difficulté. De manière cohérente, les individus R. antilleana, ayant subi l’ablation de leurs éventails, n’ont pas réussi à remonter le courant rapide, et leur performance a fortement diminué dans le courant lent. Les individus aux éventails rudimentaires ont eu une performance intermédiaire. En fait, l’analyse de la fréquence de battement montre que les animaux ayant subi une ablation de l’éventail ou à l’éventail rudimentaire tentent de compenser en battant plus de fois par seconde. Dans certain cas cela suffit, mais pas toujours.
Les chercheurs concluent ainsi que l’éventail permet d’augmenter l’efficience des battements en augmentant la surface de la patte en contact avec l’eau et l’efficacité de chaque poussée par le transfert d’une plus grande quantité d’énergie. Ainsi, moins de battements sont nécessaires en eau stagnante, et des vitesses de pointes plus rapides permettent de lutter contre le courant.
Les performances de locomotion n’ont pas été affectées par le sexe des individus.
Enfin, les scientifiques ont évalué la capacité à effectuer des virages serrés, un comportement caractéristiques des espèces de Rhagovelia. Ils ont constaté que les virages des individus normaux (témoins) étaient systématiquement initiés par le déploiement des éventails et se réalisaient avec un angle moyen de 77°. Les individus des deux groupes sans éventails (inhibition et ablation) ont quant à eux tourné de manière moins efficace avec un angle intermédiaire de 111° et 126° respectivement.
Pris ensemble, ces résultats montrent l’importance de l’éventail dans le mode de vie de Rhagovelia. Plus important encore, un éventail rudimentaire confère toujours un avantage comparé à son absence. Les chercheurs pensent ainsi u’un éventail rudimentaire est apparu à la base des Rhagovelia puis il a été graduellement amélioré par une concordance entre des changements génétiques et la sélection.
Conclusion
Les résultats montrent le lien entre des mécanismes génétiques du développement, évolutifs et écologiques lors de l’émergence d’innovations comme l’éventail.
Punaises du genre Rhagovelia antilleana à la surface de l’eau (Source : A. Khila, 2017)
L’ordre phylogénétique de ces événements moléculaires demeurent inconnu. Les chercheurs ont reconstitué la majeure partie de l’histoire évolutive de l’éventail et proposent le scénario suivant. Initialement, le gain de fonction d’anciens programmes de développement dans la seconde paire de pattes a permis l’apparition d’un éventail rudimentaire. Puis, la duplication du gène geisha et son implication dans le développement a amélioré l’éventail. Chacune des étapes a pu être sélectionnée au cours de l’adaptation à la nouvelle niche que représentaient les cours d’eau. Cette étude apporte aussi un exemple, expérimentalement démontré et clair, de comment une nouveauté évolutive peu émerger graduellement à partir de structures plus simples ; une question clé en évolution.
Source
Santos M.E., Le Bouquin A., Crumière A.J.J. & Khila A. (2017) : Taxon-restricted genes at the origin of a novel trait allowing access to a new environment. Sciences, 358:386-390 (lien)
– The Insects: Structure and Function(R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
One thought on “Evolution et adaptation chez les punaises Rhagovelia”
Bonjour,
Le 23/05/2019
Nous sommes dans le Gers à la terrasse du château de Saint Orens Pouy Petit..
Nous avons du rentrer à l’intérieur car des centaines de tout petit insectes, plus petit qu’un point sur le i, ont déclenché des démangeaisons sur nos visages
. Tous les étés au pic de la chaleur avec les cultures de blé dans la vallée en contre bas nous avons ces attaques. Les connaissez vous ?
Ce site internet utilise les cookies pour améliorer votre expérience utilisateur et à des fins statistiques et publicitaires. Les cookies nécessaires sont essentiels au bon fonctionnement du site internet. En les désactivant, cela peut nuire à votre expérience utilisateur.
Les cookies fonctionnels aident à exécuter certaines fonctionnalités telles que le partage du contenu du site Web sur les plateformes de médias sociaux, la collecte de commentaires et d'autres fonctionnalités tierces.
Les cookies de performance sont utilisés pour comprendre et analyser les indices de performance clés du site Web, ce qui contribue à offrir une meilleure expérience utilisateur aux visiteurs.
Cookie
Durée
Description
_gat
1 minute
Ce cookie est installé par Google Universal Analytics pour limiter le taux de demande et ainsi limiter la collecte de données sur les sites à fort trafic.
Les cookies analytiques sont utilisés pour comprendre comment les visiteurs interagissent avec le site Web. Ces cookies aident à fournir des informations sur le nombre de visiteurs, le taux de rebond, la source de trafic, etc.
Cookie
Durée
Description
CONSENT
2 ans
YouTube définit ce cookie via des vidéos YouTube intégrées et enregistre des données statistiques anonymes.
vuid
2 ans
Vimeo installe ce cookie pour collecter des informations de suivi en définissant un identifiant unique pour intégrer des vidéos sur le site Web.
_ga
2 ans
Le cookie _ga, installé par Google Analytics, calcule les données des visiteurs, des sessions et des campagnes et assure également le suivi de l'utilisation du site pour le rapport d'analyse du site. Le cookie stocke les informations de manière anonyme et attribue un numéro généré de manière aléatoire pour reconnaître les visiteurs uniques.
_gat_gtag_UA_48379736_2
1 minute
Défini par Google pour distinguer les utilisateurs.
_gid
1 jour
Installé par Google Analytics, le cookie _gid stocke des informations sur la façon dont les visiteurs utilisent un site Web, tout en créant un rapport d'analyse des performances du site Web. Certaines des données collectées incluent le nombre de visiteurs, leur source et les pages qu'ils visitent de manière anonyme.
Les cookies publicitaires sont utilisés pour fournir aux visiteurs des publicités et des campagnes marketing pertinentes. Ces cookies suivent les visiteurs sur les sites Web et collectent des informations pour fournir des publicités personnalisées.
Les cookies nécessaires sont absolument essentiels au bon fonctionnement du site Web. Ces cookies assurent les fonctionnalités de base et les fonctions de sécurité du site Web, de manière anonyme.
Contribuer au magazine
Vous pouvez à l'aide d'un don, quel qu'il soit, soutenir le développement du Magazine et contribuer à la sensibilisation du plus grand nombre à la préservation des insectes.











Bonjour,
Le 23/05/2019
Nous sommes dans le Gers à la terrasse du château de Saint Orens Pouy Petit..
Nous avons du rentrer à l’intérieur car des centaines de tout petit insectes, plus petit qu’un point sur le i, ont déclenché des démangeaisons sur nos visages
. Tous les étés au pic de la chaleur avec les cultures de blé dans la vallée en contre bas nous avons ces attaques. Les connaissez vous ?