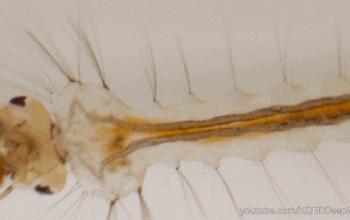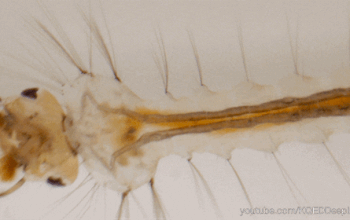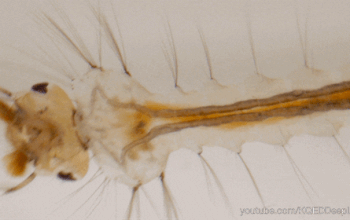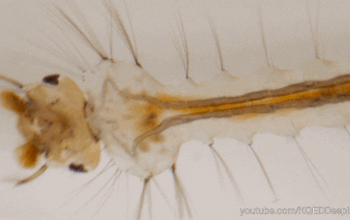
Dans l’univers des parasitoïdes (lire cet article), l’ovipositeur, également appelé oviscapte (appendice abdominal avec lequel les oeufs sont déposés), a un rôle crucial puisqu’il permet de situer précisément la ponte dans l’espace et dans le temps, ce qui est essentiel quand celle-ci doit s’accorder avec le cycle biologique de l’hôte. Avant d’aborder certaines particularités étonnantes, il faut d’abord en détailler quelque peu la structure. Chez la plupart des insectes holométaboles, l’ovipositeur est une simple excroissance tubulaire autour de l’orifice génital, mais, chez les hyménoptères, il a conservé une structure primitive bien plus complexe, qualifiée de « lépismoïde » : il s’agit en fait de deux paires de valves creuses qui forment l’ovipositeur à proprement parlé, et sont protégées par une troisième paire, le fourreau.
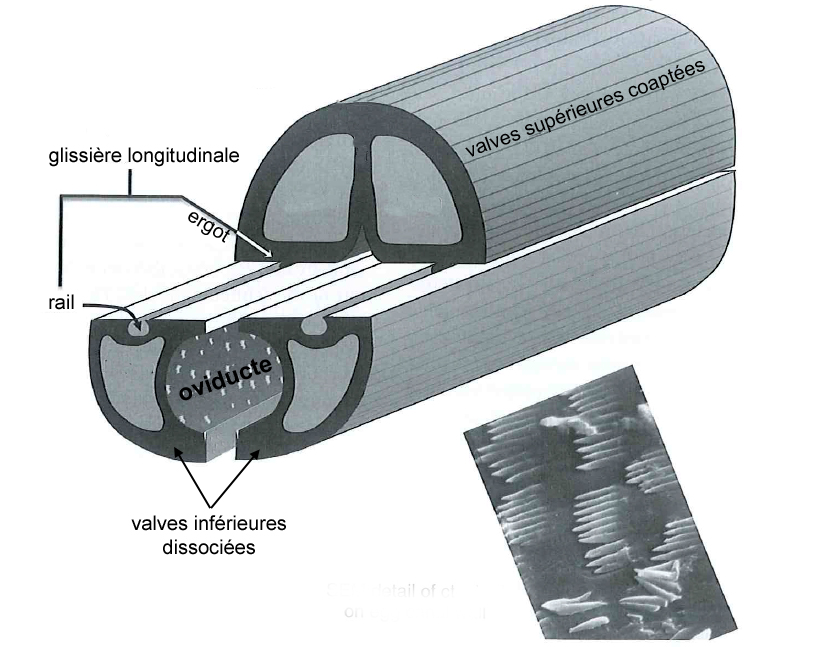
Illustration 1 : Coupe transversale d’un ovipositeur « standard » d’hyménoptère, montrant les deux paires de valves et leur articulation coulissante. En médaillon, photo MEB montrant les cténidies qui tapissent la face interne de l’oviducte et empêchent le reflux de l’oeuf lors de la ponte (Source : simplifié d’après Quicke, 2015*)
La première paire est coaptée en une pièce supérieure unique qui s’emboite dans une deuxième paire formée de deux valves inférieures, le tout formant un tube autour de l’oviducte depuis l’orifice génital (illustration 1 ci-contre). Ces deux paires sont liées par un système mécanique formant une glissière longitudinale : la face inférieure des valves supérieures forme un ergot qui s’insère dans un rail porté par les valves inférieures. Ainsi, les deux paires peuvent coulisser l’une par rapport à l’autre, ce qui joue un rôle capital lors de l’oviposition. En effet, l’ovipositeur lui-même ne contient pas de muscle : le passage de l’oeuf y est assuré à la fois par le frottement relatif des valves creuses et par la contraction des muscles abdominaux. La face interne des valves est par ailleurs dotée de structures en peignes empêchant le reflux de l’oeuf durant la ponte.
Curieusement, certains ovipositeurs ont des formes particulièrement sinueuses (Photo 1 ci-dessous). On s’est longtemps interrogé sur l’utilité d’une semblable géométrie, a priori plus gênante qu’autre chose lorsqu’il s’agit de perforer un substrat. Il est de tout de même plus facile de perforer le sol avec une aiguille qu’avec un ressort… Puis on a réalisé que le mouvement relatif des valves d’un ovipositeur sinueux a ceci d’intéressant qu’il en modifie l’orientation verticale. Ainsi, en jouant sur le frottement des deux valves, la femelle en guide l’extrémité. Vous saisissez l’utilité du dispositif lorsqu’il s’agit d’atteindre un hôte enfoui dans une galerie non rectiligne ? La recherche médicale s’est d’ailleurs empressée de récupérer cette petite merveille naturelle pour faire naviguer les instruments de microchirurgie à l’intérieur des vaisseaux sanguins.
Photo 1 : Habitus de Pristomerus hansoni – Rousse, Villemant & Seyrig. Notez la grande taille de l’ovipositeur et surtout la forme très sinueuse de son extrémité (Source : photo – Rousse / Iziko SA Museums)
La structure de base de l’ovipositeur est donc similaire chez tous les parasitoïdes. Mais cette unité structurelle est associée à une très forte diversité morphologique : la forme et la taille de l’organe varient énormément d’un groupe à l’autre et à l’intérieur même d’un groupe. La raison en est simple : la structure générale de l’ovipositeur est liée à la phylogénie, mais sa forme précise est conditionnée par l’écologie. Autrement dit, les hyménoptères, ayant tous un ancêtre commun, ont hérité d’un ovipositeur de structure analogue, mais ces hyménoptères ayant des exigences écologiques très variables, leur ovipositeur a évolué secondairement afin de s’adapter aux conditions environnementales.
Le premier trait directement lié à l’écologie du parasitoïde et à son hôte est la longueur de l’ovipositeur. Les parasitoïdes d’hôtes exposés en surface auront un ovipositeur extrêmement court, alors qu’il atteint une longueur relative impressionnante chez les parasitoïdes qui doivent fouiller de grandes profondeurs. Cette longueur relative est souvent limitée à environ 1,3 fois la taille de l’insecte, ce qui correspond grosso modo à la taille qu’il doit avoir pour que l’insecte, en extension sur ses pattes antérieures, puisse en ramener la pointe vers l’avant et diriger sa ponte. Cependant, de nombreux parasitoïdes ont développé des comportements ou des structures anatomiques qui leur permettent de dépasser cette limite. Dans des exemples extrêmes, cet ovipositeur peut ainsi faire plus de dix fois la longueur du corps de l’insecte.
Photo 2 : vues dorsale et latérale d’une femelle d’Ibalia leucospides avec son gastre en lame de couteau à l’intérieur duquel est enroulé l’ovipositeur (Source : B. Mallet, Le monde des Insectes)
Chez la plupart des parasitoïdes, l’ovipositeur est externe et pointe vers l’arrière à l’extrémité de l’abdomen (appelé également gastre). Lorsque cet ovipositeur est long, l’insecte le traine derrière lui, ce qui peut gêner son mouvement et le pénaliser face aux prédateurs. Certaines espèces à ovipositeur de grande taille ont développé au cours de l’évolution des structures de rangement que l’on imagine facilement plus confortables. Par exemple, dans la famille des Cynipidae, l’ovipositeur forme une boucle plus ou moins complète à l’intérieur de l’abdomen (photo 2), et il est complètement enroulé chez les Orussidae, des parasitoïdes de larves xylophages. Chez les Leucospidae (photo 3) et certains Platygstridae (photo 4), l’ovipositeur est dirigé vers l’avant et rangé sur la face dorsale de l’abdomen au repos.
Photo 3 : Femelle Leucospis dorsigera avec le fourreau de l’ovipositeur reposant sur la face dorsale du gastre et pointant vers l’avant (Source : D. Geystor, Le monde des Insectes)
Dans d’autres cas où l’ovipositeur est particulièrement long, le déploiement est facilité par des structures spécifiques. Ainsi, chez les Ichneumonidae du genre Megarhyssa, la partie externe de l’ovipositeur peut atteindre plus de trois fois la longueur du corps. On peut voir sur cette vidéo comment la femelle de Megarhyssa atrata procède pour insérer et retirer son ovipositeur dans un tronc dans lequel elle recherche un hôte (voir également les vidéos ci-dessous) . Cette délicate manoeuvre est assurée par la rotation complète des segments apicaux de l’abdomen couplée au déploiement d’une membrane issue de la jonction entre les segments 7 et 8, membrane elle-même associée à des glandes sécrétrices. Grâce à cet ensemble, M. atrata peut atteindre des hôtes enfouis sous 14 cm de bois dur, alors que le corps de l’animal ne mesure que 4 cm !
Photo 4 : Femelle d’Inostemma boscii avec le premier segment du gastre transformé portant un cornet courbet abritant l’ovipositeur au repos (Source : J. Svabik)
L’extrémité de l’ovipositeur est également un bon indicateur de l’écologie de son propriétaire. Les parasitoïdes ayant pour hôtes des oeufs, immobiles et fragiles, possèdent dans la majeure partie des cas un ovipositeur à l’extrémité effilée, alors qu’un hôte mobile, comme une larve vivante, requiert une extrémité en tête de flèche barbelée afin d’assurer la prise. Plus surprenante encore est la composition chimique de cet ovipositeur. Ainsi, pour les espèces dont les hôtes se trouvent dans un substrat dur comme le bois, l’ovipositeur est alors renforcé par des minéraux métalliques (manganèse ou zinc) qui en assurent la dureté !
Pascal Rousse (Source : Pascal Rousse)
Passion-Entomologie souhaite grandement remercier Pascal Rousse pour sa nouvelle contribution. Pascal est chercheur entomologiste, correspondant du Muséum National d’Histoire Naturelle de Paris (MNHN), versé dans la taxonomie, l’écologie et la phylogénie des hyménoptères parasitoïdes, en particulier dans le cadre de leur utilisation en lutte biologique, auteur d’une trentaine de publication sur ces sujets, mais avant tout fasciné depuis bien longtemps par la biologie des Ichneumonidae sans que la psychanalyse ait pu trouver une explication rationnelle à cette monomanie !
Ce site internet utilise les cookies pour améliorer votre expérience utilisateur et à des fins statistiques et publicitaires. Les cookies nécessaires sont essentiels au bon fonctionnement du site internet. En les désactivant, cela peut nuire à votre expérience utilisateur.
Les cookies fonctionnels aident à exécuter certaines fonctionnalités telles que le partage du contenu du site Web sur les plateformes de médias sociaux, la collecte de commentaires et d'autres fonctionnalités tierces.
Les cookies de performance sont utilisés pour comprendre et analyser les indices de performance clés du site Web, ce qui contribue à offrir une meilleure expérience utilisateur aux visiteurs.
Cookie
Durée
Description
_gat
1 minute
Ce cookie est installé par Google Universal Analytics pour limiter le taux de demande et ainsi limiter la collecte de données sur les sites à fort trafic.
Les cookies analytiques sont utilisés pour comprendre comment les visiteurs interagissent avec le site Web. Ces cookies aident à fournir des informations sur le nombre de visiteurs, le taux de rebond, la source de trafic, etc.
Cookie
Durée
Description
CONSENT
2 ans
YouTube définit ce cookie via des vidéos YouTube intégrées et enregistre des données statistiques anonymes.
vuid
2 ans
Vimeo installe ce cookie pour collecter des informations de suivi en définissant un identifiant unique pour intégrer des vidéos sur le site Web.
_ga
2 ans
Le cookie _ga, installé par Google Analytics, calcule les données des visiteurs, des sessions et des campagnes et assure également le suivi de l'utilisation du site pour le rapport d'analyse du site. Le cookie stocke les informations de manière anonyme et attribue un numéro généré de manière aléatoire pour reconnaître les visiteurs uniques.
_gat_gtag_UA_48379736_2
1 minute
Défini par Google pour distinguer les utilisateurs.
_gid
1 jour
Installé par Google Analytics, le cookie _gid stocke des informations sur la façon dont les visiteurs utilisent un site Web, tout en créant un rapport d'analyse des performances du site Web. Certaines des données collectées incluent le nombre de visiteurs, leur source et les pages qu'ils visitent de manière anonyme.
Les cookies publicitaires sont utilisés pour fournir aux visiteurs des publicités et des campagnes marketing pertinentes. Ces cookies suivent les visiteurs sur les sites Web et collectent des informations pour fournir des publicités personnalisées.
Les cookies nécessaires sont absolument essentiels au bon fonctionnement du site Web. Ces cookies assurent les fonctionnalités de base et les fonctions de sécurité du site Web, de manière anonyme.
Contribuer au magazine
Vous pouvez à l'aide d'un don, quel qu'il soit, soutenir le développement du Magazine et contribuer à la sensibilisation du plus grand nombre à la préservation des insectes.







)
(N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae – 750 pages – 5 septembre 2013)
(Bernard Pintureau – Editions : Ellipses Marketing – 188 pages – 25 avril 2009)