

Les yeux ont pour fonction de réceptionner et de guider l’énergie lumineuse (photons) vers des cellules réceptrices spécialisées (photorécepteurs) qui traduisent cette énergie photonique en énergie électrique. Transmise ensuite au cerveau par des neurones, elle sera analysée et interprétée pour donner naissance à une représentation mentale de l’environnement permettant à l’animal de se déplacer; de repérer sa proie ou son partenaire sexuel, etc.
Les yeux des vertébrés, constitués d’une structure unique, sont appelés yeux simples. Chez les arthropodes, les yeux sont, quant à eux, dits composés car constitués de plusieurs sous-unités similaires : les ommatidies.
Les yeux composés sont apparus tôt dans l’évolution. Ils ont été observés sur des fossiles de crustacés et de chélicérates (scorpions et araignées) du Cambrien (540-485 millions d’années) et sur des insectes du Dévonien (420-360 Ma). Cette période a été favorable à une grande diversification du monde animal. Les yeux composés de l’actuelle limule (Limulus sp., Chélicérates) sont ainsi restés inchangés depuis cette époque (lire cet article).
En raison de leur petite taille, l’observation et l’étude des yeux des insectes a du attendre l’invention du microscope au XVIIème siècle. Les premières descriptions en ont été réalisées par le chimiste, physicien et mathématicien anglais Robert Hooke en 1665, par le savant néerlandais Antoni Van Leeuwenhoek en 1695 et par le physiologiste autrichien Sigmund Exner en 1891.
I) Présence des yeux composés
Les yeux composés se retrouvent chez la quasi-totalité des espèces d’insectes : leur taille, leur forme et leur structure varient cependant entre les familles et les espèces.
Chez les Aptérygotes (dépourvus d’ailes), insectes qualifiés de « primitifs », ils sont absents par exemple chez les Thysanoures (ordre d’insectes aptères) et les Protoures (arthropodes pancrustacés hexapodes longtemps considérés comme des insectes) et ne sont constitués que de 8 ommatidies chez les autres.
Par contre, chez les Ptérygotes (pourvus d’ailes), insectes qualifiés de « modernes », le nombre d’ommatidies peut être particulièrement élevé : 800 chez les drosophiles (Diptères), 7 500 chez les Diopsidae (Diptères) (lire cet article), 10 000 chez les bourdons (Hyménoptères), ou encore 30 000 chez les Odonates (libellules).
Certaines espèces cavernicoles, souterraines comme les termites (Isoptères), parasites aux ailes atrophiées (Phthiraptera et Siphonaptera), ou de cochenilles (Hémiptères) en sont dépourvus. L’oeil de la fourmi Hyponera punctatissima (Hyménorptères – Formicidae) n’est constitué, quant à lui, que d’une seule ommatidie.
II) Structure d’une ommatidie
Chaque ommatidie est constituée de 3 parties : optique, de collecte de la lumière et sensorielle.
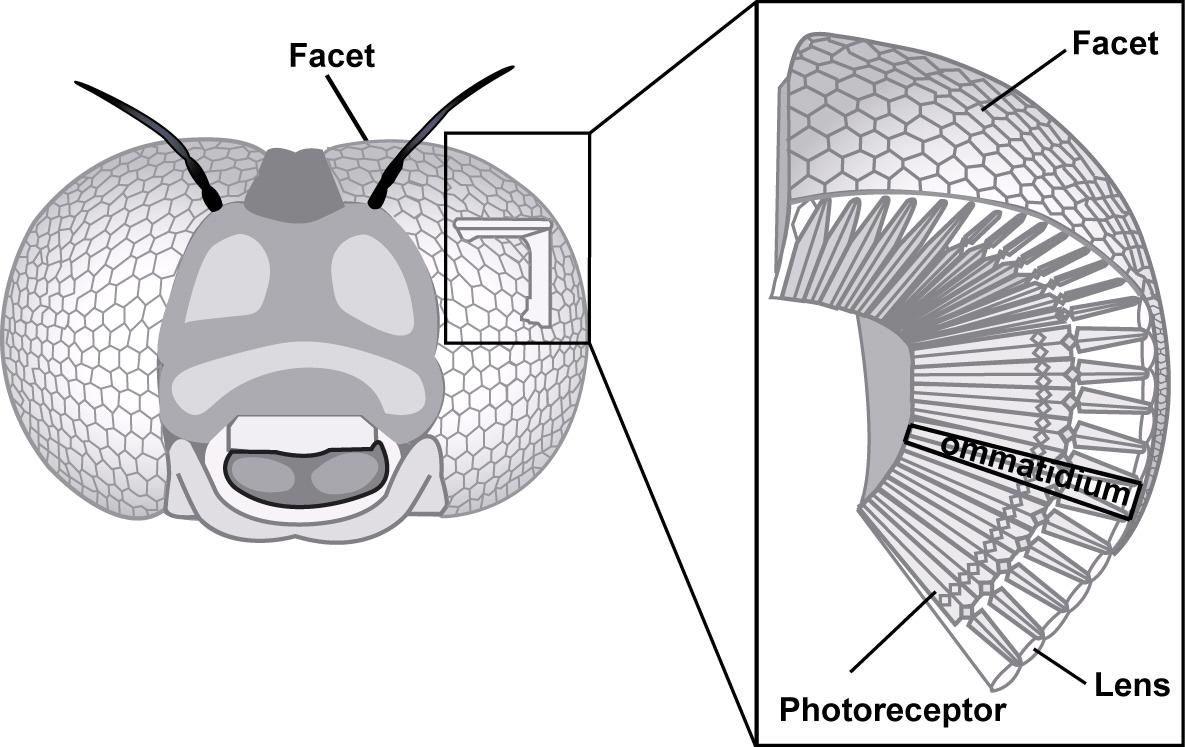
Comme l’ensemble du corps des insectes, les yeux composés sont couverts d’une cuticule (lire cet article), mais pour des raisons évidentes, celle-ci est à cet endroit transparente, incolore et forme une cornée, ou lentille biconvexe, à la surface de chaque ommatidie.
Vu de dessus, les ommatidies forment des facettes hexagonales placées les unes à coté des autres (figure 1).
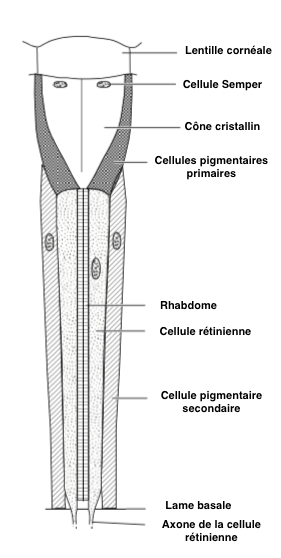
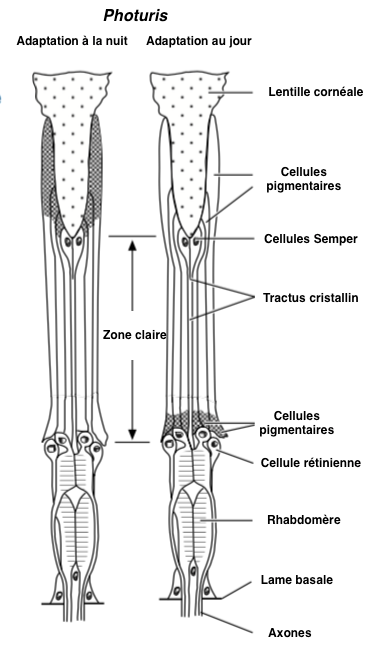
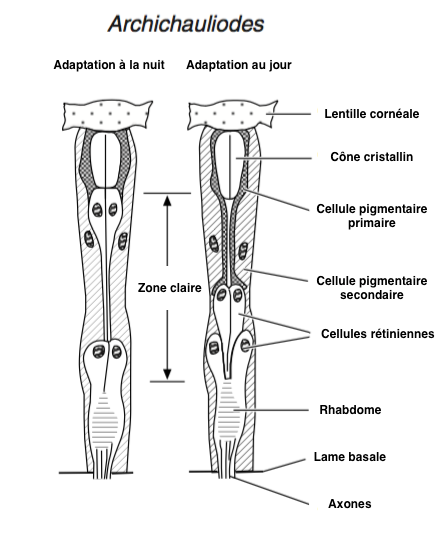
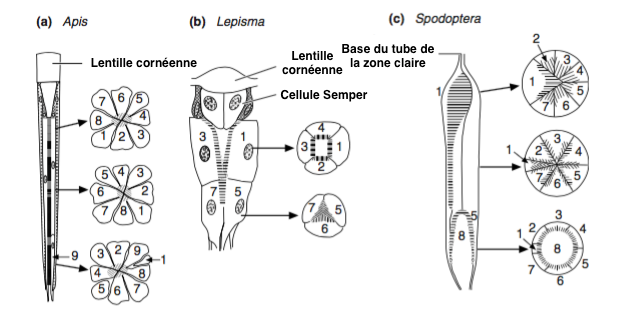
La cornée est synthétisée et sécrétée par deux cellules épidermiques, cellules cornéales, ou aussi cellules pigmentaires primaires. Sous la cornée se trouvent 4 cellules (cellules Semper) dont la fonction est de produire, chez certaines espèces, une seconde lentille rigide et transparente appelée cône cristallin (Figure 2). Sous la Cornée et le cône cristallin sont placées les cellules sensorielles (photorécepteurs ou cellules rétiniennes) qui sont des neurones allongés, généralement au nombre de 8, parfois de 7 ou de 9, bordées latéralement par 12 à 18 cellules de soutien, séparant les ommatidies entre elles, et appelées cellules pigmentaires secondaires (figure 2). Les photorécepteurs traversent la lame basale (lamina basale) où ils se connectent aux neurones du nerfs optique au niveau du lobe optique.
La marge interne des photorécepteurs est composée de microvillosités très denses et perpendiculaires à l’axe de la cellule : le rhabdomère, où se situent les pigments visuels comme la rhodopsine.
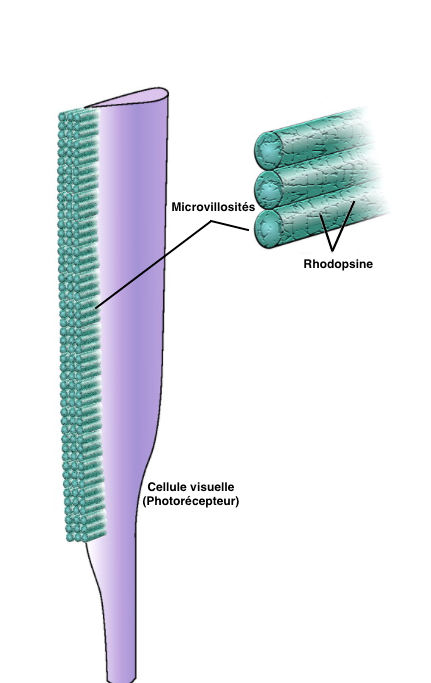
L’union des microvillosités de tous les photorécepteurs forme le rhabdome (voir figure 3).
Chez certaines espèces d’abeilles ou de mouches, les photorécepteurs réalisent une torsion sur leur longueur modifiant ainsi l’orientation des microvillosités ce qui a pour effet d’éliminer les effets de la polarisation (explication lors d’un prochain article).
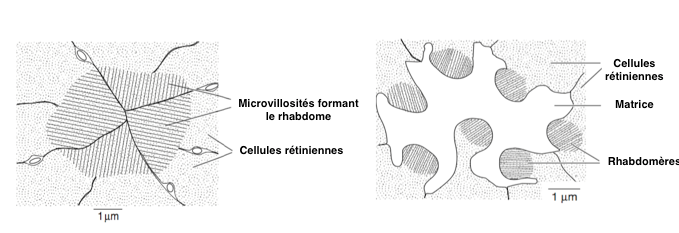
Les rhabdomères peuvent être soit » fusionnés », soit « ouverts ». Dans le premier cas, tous les rhabdomes sont en contact les uns avec les autres et ont le même champ visuel. Dans le second, ils sont séparés par une matrice et ont chacun un champ visuel propre (voir figure 4).
La configuration des yeux composés diffère si les insectes sont diurnes ou nocturnes. La quantité de lumière étant plus faible la nuit, l’évolution a modifié la structure des ommatidies et des yeux composés pour augmenter l’efficacité de capture des photons.
- Structure en apposition : elle se rencontre principalement chez les espèces diurnes. Le rhabdome s’étend sur toute la longueur de la cellule photoréceptrice, du cône cristallin à la lame basale, soit une longueur de 150 µm chez les fourmis Camponotus et 80 µm chez la mouche drosophile (voir figure 5A). Les photorécepteurs ne reçoivent la lumière que de la lentille à laquelle ils sont liés.
- Structure en superposition : elle se rencontre principalement chez les espèces nocturnes (papillons et coléoptères). Le rhabdome ne s’étend pas sur toute la longueur de la cellule photoréceptrice (voir figure 5B). Les photorécepteurs reçoivent la lumière de centaines, voire de milliers, de facettes augmentant la capture des photons et donc la sensibilité visuelle.

Remarques : les Hyménoptères (guêpes, abeilles, fourmis) possèdent des yeux de type apposition. Bien que ce type de vision offre une faible sensibilité lumineuse, certaines espèces, devenues nocturnes au cours de l’évolution, possèdent contre toute attente une bonne acuité visuelle. De nombreux scientifiques, comme l’australien Eric J. Warrant (2008), mènent des expérimentations afin de percer ce mystère.
Les 12 à 18 cellules pigmentaires secondaires isolent les ommatidies les unes des autres.
Une autre adaptation à la vie nocturne consiste, chez les papillons notamment, en l’aplatissement et en l’élargissement des trachées respiratoires à l’arrière de l’oeil pour former une couche appelée tapetum. Cette structure a pour fonction de refléter la lumière. Les photons traversent ainsi deux fois les rhabdomes, augmentant le nombre de pigments visuels excités et donc la sensibilité de la vision.
Une vidéo décrivant tout cela sous forme d’animation :
III) Adaptation externe des ommatidies
La forme et l’arrangement des ommatidies différent entre les espèces d’insectes.
Par exemple, chez les mantes-religieuses (Mantoptères), la capture des proies nécessite une vision binoculaire à haute résolution. Pour cela, l’évolution les a doté d’yeux composés où le diamètre des facettes, l’angle interommatidal et la longueur des rhabdomes des ommatidies sont plus importants au centre pour diminuer vers les périphéries (voir figure 6). Ces caractéristiques offrent à l’insecte une zone visuelle de grande précision appelée fovéa (voir infographie ci-dessous). Ces adaptations se retrouvent également dans les yeux des vertébrés.


La zone de haute résolution visuelle se situe à l’avant des yeux composés chez les mantes-religieuses (infographie ci-contre), sur la partie fronto-dorsale des yeux composés des libellules (Odonates) qui chassent et poursuivent des proies aériennes. D’autres insectes, comme les Coléoptères du genre Gyrinus (famille des Gyrinidae) qui nagent à la surface de l’eau, possèdent deux zones de haute résolution visuelle, l’une sous la surface (ventrale) et la seconde aérienne (dorsale). Plus surprenant encore, les yeux composés des mâles du genre Cloeon (Ephéméroptères) sont constitués de deux parties indépendantes. L’une, dorsale, est constituée d’ommatidies à larges facettes et de type superposition et l’autre, latérale, à de facettes plus petites et de type apposition (voir album photos ci-dessous).
IV) Adaptation interne des ommatidies
La forme du cristallin est variable entre les espèces d’insectes. Il est en général conique, hormis pour les insectes dits « primitifs » comme les collemboles et les Thysanoures chez lesquels il est sphérique.
Il a été décrit 4 types de cône cristallin :
- Eucône : les cellules Semper se trouvent dans le cône cristallin (figure 7)
- Acône : absence de cône cristallin, le rôle de la mise au point est joué par les cellules Semper dont le cytoplasme est clair. Cette configuration se rencontre chez les Coccinellidae, les Staphylinidae, les Tenebrionidae, certains Diptères et Hémiptères
- Pseudocône : les cellules Semper sécrètent un cône cristallin extracellulaire formé de liquide gélatineux (certains Odonates, Coléoptères et la plupart des Diptères)
- Exocône : les cellules Semper ne contribuant pas à la mise au point, c’est la lentille cornéenne qui, en formant un cône, joue ce rôle (Elateridae et Lampyridae) (Figure 8)
Les photorécepteurs sont en général de même longueur. Cependant, chez des espèces d’insectes dites « primitives » comme le lépisme (Thysanoures) et les papillons du genre Spodoptera, les photorécepteurs sont enroulés et disposés de manière étagée. D’autres configurations sont également possibles (figure 9).
Pour ceux qui souhaitent poursuivre, retrouvez le deuxième article qui est consacré à la perception des formes, des distances et de la vitesse. Suivre ce lien.
Source :
– The Insects : Structure & Function (5ème édition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
– Encyclopedia of Insects : Vincent H. RESH & Ring T. CARDE, Academic Press-2009
– Srinivasan M. V. (2011) : Honeybees as a model for the study of visually guided flight, navigation and biologically inspired robotics – American Physiological Society, Vol.91 (2) 413-460 (lien)
– Warrant E. J. (2008) : Seeing in the dark : vision and visual behavior in nocturnal bees and wasps – Journal of Experimental Biology, 211:1737-1746 (lien)
Recommandation d’ouvrages sur cette thématique
– Evolution of the Insects (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
– The Insects: An Outline of Entomology (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
– The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
– Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
{kind=link}
Merci pour cette « leçon de choses »
Comme d’habitude nous en apprenons toujours plus sur la vie qui nous entoure.
Bonne journée