L’ANR DISLAND : un projet de génétique du paysage pour informer la dispersion de la mouche orientale des fruits dans les paysages agricoles sénégalais
Par Marie-Pierre Chapuis & Karine Berthier
La mouche orientale des fruits, un ravageur majeur dans le monde
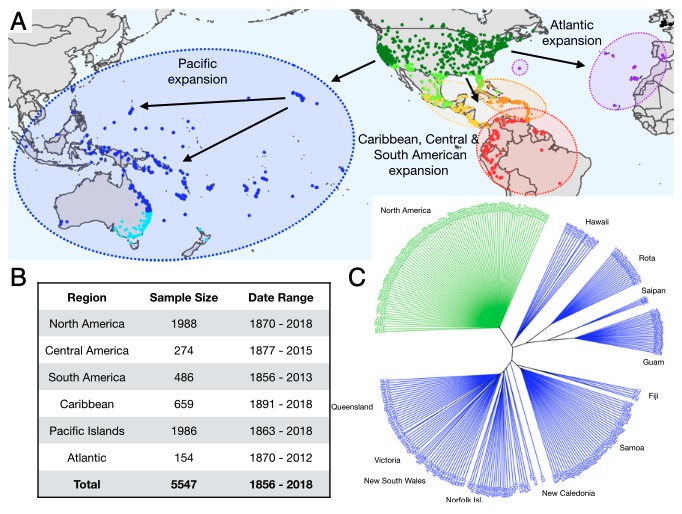
Bactrocera dorsalis (BD ; figure 1A) est une mouche (Diptera : Tephritidae), originaire de l’Asie tropicale et subtropicale. Si elle a été signalée pour la première fois en dehors de son aire native, à Hawaï, en 1942, c’est le début du siècle actuel, période d’un accroissement important des flux commerciaux internationaux, qui marque le début d’une invasion à l’échelle globale dont la grande majorité du continent africain et des îles de l’Océan Indien (figure 1B). Très polyphage, avec plus de 300 plantes hôtes recensées, elle s’est alors imposée comme un ravageur de nombreuses espèces cultivées de fruits et légumes tropicaux et subtropicaux.

La femelle pond sous la surface du fruit et les larves se nourrissent de la pulpe, le rendant impropre à la consommation. Les pertes de rendement sont considérables pour les producteurs, parfois jusqu’à la totalité d’un bassin de production, notamment sur son hôte principal, la mangue (Mangifera indica) (figure 2A). Classé organisme de quarantaine prioritaire aux États-Unis d’Amérique, en Australie et dans l’Union Européenne, BD cause des pertes économiques supplémentaires du fait de la perte d’accès à certains marchés d’export et de la destruction des lots contaminés aux frais des exportateurs ou des producteurs.
La limitation des dégâts liés à la mouche orientale des fruits est ainsi un enjeu crucial pour la filière mangue dans les régions du monde où elle est présente.
Actuellement, pour lutter contre ce fléau en Afrique de l’Ouest, les producteurs recourent toujours à l’utilisation de pesticides largement remise en cause du fait de l’apparition de résistances, des risques pour la santé humaine et de leurs coûts écologiques et économiques. Les techniques respectueuses de l’environnement employées se limitent à l’assainissement des vergers (par ex., ramassage des fruits infestés qui sont ensuite enfouis ou brûlés) (figure 2C) et à l’utilisation de méthodes d’attraction et de destruction ou de piégeage de masse.
Le CIRAD et ses partenaires sénégalais (l’Institut Sénégalais de Recherche Agricole et l’Université Cheikh Anta Diop de Dakar) développent depuis plusieurs années des techniques de gestion biologique (figure 2B), tels que la facilitation d’auxiliaires naturels (par ex. la fourmi prédatrice Oecophylla longinoda – figure 2D), le lâcher d’auxiliaires commercialisés (par ex. le parasitoïde micro-hyménoptère Fopius arisanus) et l’entomovectorisation, qui consiste à utiliser des mâles stériles pour infecter les populations sauvages de BD avec un champignon entomopathogène (Projet ANR LEAP Agri PFF – Pest Free Fruit).
L’efficacité de l’ensemble de ces techniques reste toutefois insuffisante si elles sont utilisées de manière curative à l’échelle de la parcelle agricole.

Une connaissance insuffisante de la dynamique des populations
L’abondance d’une population de ravageur dans une parcelle donnée ne dépend pas uniquement des caractéristiques agronomiques de cette parcelle (par ex. variétés cultivées) ou des capacités biologiques des individus qui la composent (par ex. fécondité, dispersion, etc), mais également des interactions étroites entre cette population et les caractéristiques de l’agro-écosystème dans lequel elle se développe.
La gestion de la mouche orientale des fruits doit ainsi être envisagée à une échelle cohérente avec les processus écologiques à l’œuvre dans les territoires concernés. Or, les effets de l’environnement sur la dynamique des populations de BD restent aujourd’hui incompris.
Dans de nombreuses régions du monde, la mouche présente une fluctuation annuelle importante de l’abondance de ses populations. La dynamique est caractérisée par : 1) un pic d’abondance qui coïncide avec la saison des pluies, la période de maturation et de récolte des mangues, et 2) une forte diminution de l’abondance, voire une disparition presque complète des populations, qui coïncide avec la saison sèche et la période d’absence des mangues.
Cette dynamique pose la question de la disponibilité et de la qualité d’habitats qui permettent à des populations (relictuelles) de persister en dehors de la saison de production (réservoirs), et, ainsi, de recoloniser des vergers en tout début de la saison de production de mangues (corridors).
Ces informations sur le fonctionnement démographique des populations peuvent théoriquement être obtenues à partir d’études utilisant des techniques de capture-marquage-recapture ou de suivis d’abondance par piégeage. Cependant, la première technique, en plus d’être d’extrêmement coûteuse, est très complexe à mettre en place pour des insectes ravageurs de cultures du fait de leur petite taille, de leur forte mortalité et des densités importantes de leurs populations.
Les suivis d’abondance, quant à eux plus simples à mettre en place, ne donnent aucune indication sur la dispersion. Une alternative à ces approches dites «directes» consiste à mieux comprendre le fonctionnement démographique des populations dans les agroécosystèmes à partir de données génétiques neutres, dans le cadre d’applications en génétique dite du paysage.
Une approche novatrice de génétique du paysage « agroécologique »
Une alternative à ces approches dites «directes» consiste à mieux comprendre le fonctionnement démographique des populations dans les agroécosystèmes à partir de données génétiques neutres, dans le cadre d’applications en génétique du paysage.
Les approches de génétique du paysage combinent les concepts et les données de la génétique des populations, de l’écologie du paysage et des statistiques spatiales. Cette discipline a émergé dans un contexte de biologie de la conservation pour caractériser l’influence du paysage sur la structure génétique des populations, afin d’évaluer la connectivité entre ces populations et de permettre la mise en place de mesures de gestion appropriées. Les espèces étudiées sont généralement inféodées à des habitats naturels distribués dans une matrice paysagère hostile et caractérisées par des populations qui ont des petites tailles, des taux de dispersion faibles et des cycles de vie assez longs.
En revanche, les populations de ravageurs se développent dans des paysages agricoles souvent très hétérogènes dans le temps et l’espace, du fait notamment des pratiques agricoles et de la saisonnalité des cultures. Elles sont également souvent caractérisées par de fortes abondances, des taux de dispersion élevés et des cycles de vie rapides : autant de facteurs qui limitent le degré de différenciation génétique et masquent les liens entre structure génétique et structure du paysage. De fait, l’application des outils de génétique du paysage reste aujourd’hui très limitée pour les ravageurs.
Le projet ANR DISLAND propose d’adapter, de développer et ainsi d’étendre le cadre de la génétique du paysage pour caractériser les dynamiques de population de ravageurs de cultures dans un contexte finalisé de gestion intégrée.
Nous relevons des défis liés notamment à 1) l’effort d’échantillonnage, à la fois en termes d’individus dans le paysage et de marqueurs dans le génome, nécessaire pour caractériser les changements rapides de la structuration génétique spatiale des populations de ravageurs ; 2) la multiplicité et la volatilité des facteurs de variabilité spatiale et temporelle des paysages agricoles ; 3) la difficulté de développement de modèles spatialisés réalistes pour estimer les processus démographiques (taille de population, dispersion).
Défis d’acquisition des données démo-génétiques sur le ravageur
La génétique du paysage se démarque de la génétique des populations par la nécessité de collecter des échantillons individuels référencés par des coordonnées géographiques (sans les regrouper a priori en populations), et cela en grand nombre de manière à maximiser la couverture de l’espace d’étude.
De plus, alors que dans les faits les études de génétique du paysage sont généralement des instantanés, il semble nécessaire de mettre en œuvre des échantillonnages longitudinaux pour représenter la dynamique saisonnière des interactions entre les populations du ravageur et l’environnement dans lequel elles s’insèrent.
De même, bien que rarement entreprises, les approches examinant pour un même ravageur plusieurs bassins de production sont nécessaires pour révéler des effets environnementaux sur la dynamique des populations qui peuvent être spécifiques de certains contextes paysagers. Enfin, il est rare de considérer conjointement une échelle spatiale supérieure à celle du bassin de production, alors que la dispersion passive à longue distance, du fait des activités humaines ou du vent, est avérée au moins chez les micro-organismes et les insectes, principaux ravageurs des cultures.
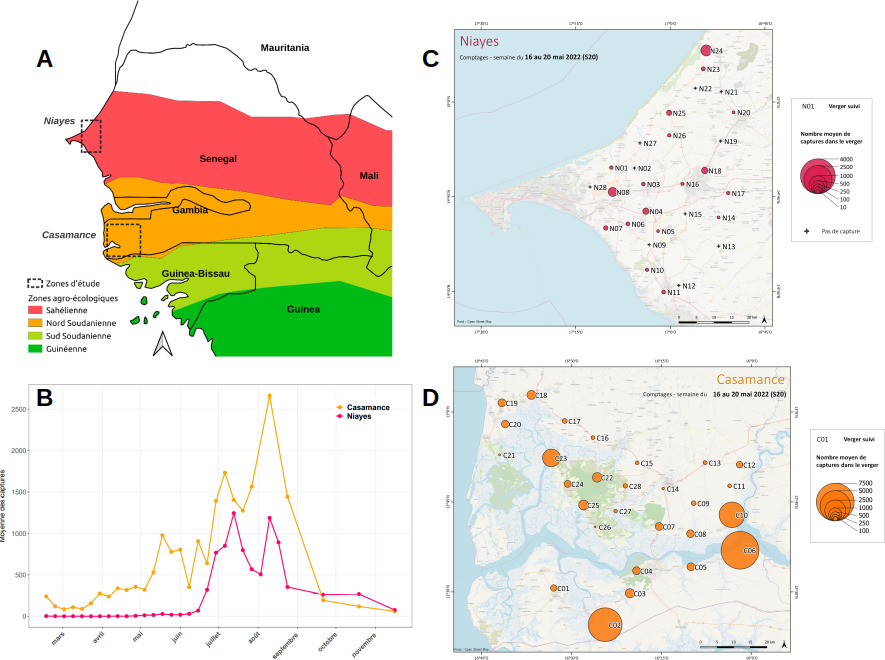
Fort d’un ancrage au Sénégal, nous avons mis en place un dispositif d’échantillonnage spatio-temporel d’envergure, comprenant un total de 56 vergers et 38 relevés annuels pour deux années successives. Les vergers sont répartis dans deux bassins de production de mangues (> 2000km2 par bassin) situés à ~ 200km de distance, l’un à proximité de Dakar, les Niayes, et l’autre au sud du Sénégal, la Basse-Casamance (figure 3). Ce dispositif permet ainsi à la fois de considérer conjointement les échelles de dispersion à courte et à longue distances et également d’étudier des paysages aux caractéristiques contrastées.
Le dispositif est caractéristique des piégeages de masse effectués sur les ravageurs des cultures, lors des suivis d’abondance ou comme technique mécanique de contrôle, et il pourrait être mis en place sans grande difficulté pour d’autres espèces. Les vergers sont répartis dans deux bassins de production de mangues (> 2000km2 par bassin) situés à ~ 200km de distance, l’un à proximité de Dakar, les Niayes et l’autre au sud du Sénégal, la Basse-Casamance (Figure 3A). Ce dispositif permet ainsi à la fois de considérer conjointement les échelles de dispersion à courte et à longue distances et également d’étudier des paysages aux caractéristiques contrastées.
Nous utilisons une technique de piégeage de masse effectuée en routine pour le contrôle mécanique de l’espèce ou déployée pour d’autres suivis d’abondance. Des pièges géo-référencés sont appâtés au méthyl-eugénol, une pro-hormone pour la phéromone sexuelle mâle de l’espèce qui a un pouvoir d’attraction spécifique sur environ 500 m (figure 4A). Au-delà des suivis d’abondance (figure 4B-C), plusieurs milliers d’échantillons individuels de BD collectés au cours du suivi pluriannuel sont typés par séquençage haut débit à l’aide de plusieurs dizaines de milliers de marqueurs génétiques, choisis pour leur qualité et ciblés par la technique, peu onéreuse, de RAD-Capture (figure 4D).
Nos stratégies d’échantillonnage et de génotypage devraient permettre de décrire précisément la variation spatio-temporelle de la diversité génétique et des flux de gènes au sein et entre les deux bassins de production sénégalais. Ce niveau de résolution fait actuellement défaut dans l’étude des insectes ravageurs.

Défis d’acquisition des données paysagères du système agroécologique
Les méthodes de génétique du paysage sont principalement dédiées à l’identification d’obstacles physiques à la dispersion, tels que les routes, les chaînes de montagnes ou les régions agricoles, dans le but de définir des unités de gestion ou de restaurer des corridors de dispersion. Par conséquent, ces études utilisent généralement des données environnementales relativement génériques et facilement accessibles, telles que des typologies d’occupation du sol à grande échelle ou des modèles numériques de terrain.
Dans le cas des ravageurs de cultures, ces informations sont souvent insuffisantes voire non significatives au regard des systèmes agroécologiques étudiés.
En effet, les paramètres démographiques clés pour l’optimisation des stratégies de gestion dépendent de nombreuses composantes, à la fois biotiques (par ex. diversité des plantes hôtes, ennemis naturels) et abiotiques (par ex. pratiques agricoles, climat), qui, en outre, exercent une influence à diverses échelles du paysage agricole.
Cette matrice environnementale complexe peut aussi être très hétérogène, spatialement et temporellement, du fait notamment de la saisonnalité des cultures et des systèmes sociotechniques. Par conséquent, la discipline peine à acquérir les données paysagères pertinentes pour comprendre la dynamique des populations de ravageurs et à les intégrer à une échelle cohérente avec leurs effets.
La complémentarité des partenaires de notre collectif de recherche (par ex. modélisation, télédétection, informatique, ingénierie des systèmes sociotechniques) nous permet d’acquérir de nombreuses données aux échelles : 1) du verger (figure 5A-B), au travers de relevés de composition en plantes hôtes, de suivis phénologiques des plantes hôtes, d’enquêtes auprès des producteurs sur les pratiques agricoles et sur les pertes de récoltes, et d’estimations des dégâts sur les fruits, 2) du bassin de production, au travers d’approches de télédétection qui classifient les éléments du paysage (figure 5C) en termes de favorabilité pour la mouche des fruits, et 3) de la région, au travers, d’une part, d’enquêtes auprès des filières commerciales de transport de mangues potentiellement contaminées à partir des bassins de production plus précoces (par ex. Casamance, Guinée, Guinée-Bissau et Mali), d’autre part, de modèles climatiques (par ex. températures, précipitations et humidité relative du sol ou de la canopée) ou de reconstruction de trajectoires de masses d’air pouvant entraîner des mouches sur de longues distances.

Défis de développement de modèles spatialement explicites
Les approches d’estimation de paramètres démographiques à partir de données génétiques ont historiquement été développées dans des contextes non-spatialisés, peu réalistes aux échelles de nos études et pour les espèces considérées. Dans ce contexte, notre projet a pour ambition de mettre en œuvre deux méthodologies différentes d’estimation de paramètres démographiques dans un contexte spatialisé, qui toutes deux reposent sur l’utilisation de développements statistiques récents et performants.
L’une est basée sur les modèles d’isolement par la distance de la génétique des populations, tandis que l’autre s’appuie sur les modèles de diffusion de la dynamique des populations et de l’épidémiologie. Cette dernière permet d’estimer les paramètres démographiques à partir de données diverses (génétiques, mais également comptages, paysage) ne reflétant qu’une observation partielle des processus étudiés. Elle est aussi plus flexible pour intégrer l’hétérogénéité spatiale et temporelle de l’environnement sur les paramètres démographiques que la méthodologie issue de la génétique des populations. En effet, celle-ci nécessite une définition préalable des zones d’hétérogénéité des paramètres démographiques, information que nous fournirons au travers d’analyses descriptives de la variation génétique.
Autour d’un même questionnement et de données communes, nous serons en mesure d’évaluer les performances et limites de ces deux approches, leurs différences et leurs complémentarités, leur faisabilité et leur intérêt pour la gestion.
Nous nous demanderons en particulier : à quels paramètres démographiques d’intérêt en lien avec le paysage peut-on avoir accès, et avec quelle précision ? Quel niveau de complexité peut-on envisager dans la modélisation de l’hétérogénéité du paysage ? Quel est le compromis pertinent entre précision des estimations des paramètres d’intérêt et coût de production des données génomiques ?
Ainsi, nous pourrons proposer des recommandations sur les stratégies d’échantillonnage, définir le périmètre d’application des méthodes et réorienter les efforts de recherche dans ce domaine. Éventuellement, nous pourrons proposer de nouvelles solutions ‘couplées’ pour réconcilier les approches génétiques et démographiques.
Une évolution vers une gestion intégrée et optimisée
La compréhension des processus de reproduction, de survie et de dispersion qui conditionnent l’infestation saisonnière des cultures devrait ouvrir des opportunités pour faire évoluer les pratiques de gestion, basées sur des décisions individuelles à l’échelle du champ cultivé, vers une organisation collective et une coordination des actions à l’échelle des territoires.
Par exemple, il deviendrait possible de cibler les populations aux échelles pertinentes qu’elles soient spatiales (par ex., le bassin de production) ou temporelles (par ex. saison sèche), en s’appuyant également sur la surveillance pour aider à la prise de décision, sur la base de seuils d’intervention.
La mise en œuvre de telles interventions préventives permettrait tout à la fois de réduire les dégâts et l’application des pesticides. Également, d’autres techniques de gestion pourraient être utilisées en combinaison avec les agents de lutte biologique et la gestion physique par piégeage de masse, comme la gestion des cultures (par ex., irrigation) et la manipulation du paysage (par ex., disposition spatiale des cultures hôtes).
De plus, si cette connaissance sur le système écologique (ravageur et paysage agricole) est un prérequis pour le succès d’une stratégie de gestion intégrée, elle reste insuffisante.
En effet, les innovations sont rarement adoptées si elles ne sont pas compatibles avec les systèmes sociotechniques dominants (par ex. freins sociotechniques aux innovations). Nous avons établi une forte coopération avec les acteurs des systèmes sociotechniques concernés, notamment les producteurs de mangues au Sénégal, afin de transférer les résultats de notre recherche, prendre en compte leurs besoins et contraintes et in fine accompagner la transition vers une stratégie de gestion durable et optimisée de la mouche orientale des fruits.
Le collectif de l’ANR Disland
Pour répondre à la problématique de l’ANR, nous avons constitué un collectif interdisciplinaire composé de techniciens, ingénieurs et chercheurs du CIRAD, de l’INRAe et de l’IRD, travaillant tous sur des thématiques écologiques mais aux interfaces de l’agronomie, l’épidémiologie, la génétique des populations, la dynamique des populations, la modélisation spatiale, la télédétection, ou encore de l’ingénierie des systèmes socio-techniques (Figure 6).
Si l’ANR est coordonnée depuis Montpellier, au Centre de Biologie et de Gestion des Populations (CBGP), le collectif est réparti entre la France, notamment Montpellier (unités AIDA, HortSys, TETIS), Avignon (Pathologie Végétale et BioSP), et Gif-sur-Yvette (EGCE) et le Sénégal, avec l’Institut Sénégalais de Recherche Agricole (ISRA) et l’Université de Ziguinchor.

Les autrices
 Karine Berthier développe des approches de génétique du paysage afin d’identifier et de hiérarchiser les facteurs environnementaux, y compris socio-économiques, dont la variabilité, spatiale et temporelle, influence les processus démographiques et évolutifs dans les populations de pathogènes et de ravageurs de cultures.
Karine Berthier développe des approches de génétique du paysage afin d’identifier et de hiérarchiser les facteurs environnementaux, y compris socio-économiques, dont la variabilité, spatiale et temporelle, influence les processus démographiques et évolutifs dans les populations de pathogènes et de ravageurs de cultures.
 Marie-Pierre Chapuis s’intéresse à l’écologie moléculaire et évolutive des populations d’insectes ravageurs de cultures, pour déduire leur histoire évolutive (par ex., origine géographique des populations, reconstruction des routes d’invasion ou de colonisation) ou leur dynamique contemporaine (par ex., la dispersion). Ce travail peut inclure une révision systématique des taxons étudiés, la caractérisation de facteurs environnementaux (par ex., paysage ou le microbiote intestinal), et le développement de méthodes d’analyse moléculaire ou statistique (lien).
Marie-Pierre Chapuis s’intéresse à l’écologie moléculaire et évolutive des populations d’insectes ravageurs de cultures, pour déduire leur histoire évolutive (par ex., origine géographique des populations, reconstruction des routes d’invasion ou de colonisation) ou leur dynamique contemporaine (par ex., la dispersion). Ce travail peut inclure une révision systématique des taxons étudiés, la caractérisation de facteurs environnementaux (par ex., paysage ou le microbiote intestinal), et le développement de méthodes d’analyse moléculaire ou statistique (lien).
L’agroécologie vous intéresse ? Vous souhaitez mettre en place des études, des mesures et des suivis au sein de vos parcelles et de votre exploitation : https://www.solinbio.fr
Bibliographie
- Ali OA ; O’Rourke SM ; Amish SJ ; Meek MH ; Luikart G ; Jeffres C & Miller MR (2016) : RAD Capture (Rapture) : Flexible and Efficient Sequence-Based Genotyping. Genetics 202(2): 389-400 (lien)
- Balkenhol N ; Cushman S ; Storfer A & Waits L (2015) : Landscape Genetics: Concepts, Methods, Applications. John Wiley & Sons, Chichester, UK. (lien)
- Brévault T & Clouvel P (2018) : Pest management : Reconciling farming practices and natural regulations. Crop Protection 115: 1-6 (lien)
- Caumette C ; Diatta P ; Piry S ; Faye E ; Chapuis MP & Berthier K (2021) : Facteurs de ré-infestation des vergers par la mouche des fruits Bactrocera dorsalis dans le bassin de production horticole des Niayes au Sénégal. Poster. 3ème Conférence sur l’intensification durable. Dakar, Sénégal, 23-26 novembre 2021 (lien)
- Chapuis M-P ; Popple J-A ; Berthier K ; Simpson SJ ; Deveson T ; Spurgin P ; Steinbauer MJ & Sword GA (2011) : Challenges to assessing connectivity between massive populations of the Australian Plague locust. Proceedings of the Royal Society of London B 278(1721): 3152-3160 (lien)
- Correa P ; Wäckers F ; Brévault T ; Bouvery F ; Detrain C & Chailleux A (2022) : Sugar feeders reduce weaver ants’ drawbacks when used as biological control agents in mango orchards. Biological Control 177 :105103 (lien)
- Diop S ; Dosso F ; Brévault T & Chailleux A (2021) : Contrôle des populations de mouches des fruits par dissémination assistée d’un champignon entomopathogène. Poster. 3ème Conférence sur l’intensification durable. Dakar, Sénégal, 23-26 novembre 2021 (lien)
- Drew R ; Tsuruta K & White I (2005) : A new species of pest fruit fly (Diptera: Tephritidae: Dacinae) from Sri Lanka and Africa. African Entomology 13 : 149–154 (lien)
- Ndiaye D ; Brévault T & Belmin R (2019) : Freins à la diffusion des technologies de lutte biologique contre la mouche des fruits Bactrocera dorsalis. Poster. 2ème Conférence intensification durable. Dakar, Sénégal, 8-10 octobre 2019 (lien)
- Ndlela S ; Niassy S & Mohamed SA (2022) : Important alien and potential native invasive insect pests of key fruit trees in Sub-Saharan Africa : Advances in sustainable pre- and post-harvest management approaches. CABI Agriculture and Bioscience 3(1) :7 (lien)
- Papaïx J ; Soubeyrand S ; Bonnefon O ; Walker E ; Louvrier J ; Klein E & Roques L (2022) : Inferring Mechanistic Models in Spatial Ecology Using a Mechanistic-Statistical Approach. In Statistical Approaches for Hidden Variables in Ecology (eds N. Peyrard and O. Gimenez) (lien)
- Piry S ; Chapuis M-P ; Gauffre B ; Papaïx J ; Cruaud A & Berthier K (2016) : Mapping Averaged Pairwise Information (MAPI): A new exploratory tool to uncover spatial structure. Methods in Ecology and Evolution 7(12): 1463-1475 (lien)
- Schutze MK ; Mahmood K ; Pavasovic A ; Bo W, Newman J ; Clarke AR ; Krosch MN & Cameron SL (2015) : One and the same: Integrative taxonomic evidence that Bactrocera invadens (Diptera: Tephritidae) is the same species as the Oriental fruit fly Bactrocera dorsalis. Systematic Entomology 40(2): 472–486 (lien)
- Virgoulay T ; Rousset F & Leblois R (2021) : GSpace: an exact coalescence simulator of recombining genomes under isolation by distance. Bioinformatics 37(20): 3–3675 (lien)