Le contrôle d’altitude chez les abeilles
Par Julien Serres
-
Comment les abeilles perçoivent-elles leur altitude ?
Tout comme un aviateur pilotant à vue, les apidés se servent de leur vision pour ajuster leur altitude. Les abeilles mellifères (Apis mellifera) et les bourdons sont connus pour être particulièrement sensibles au défilement optique du sol, appelé aussi flux optique, pour ajuster leur altitude en maintenant constante cette vitesse optique de défilement du sol pendant leur vol de croisière en suivant le relief d’un terrain (Portelli et al., 2010 ; Srinivasan, 2011 ; Lecoeur et al., 2019), sachant que, typiquement, la vitesse de croisière des abeilles domestiques en vol extérieur est d’environ 8 m/s (von Frisch, 1993, page 189), et que la hauteur de vol des apidés au-dessus du sol est d’environ de 2 m (Riley & Osborne, 2001).
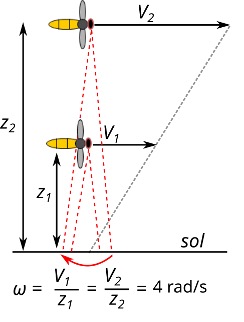
Dans leurs trajets quotidiens pour fourrager (activité de collecte du nectar et du pollen), on peut estimer que les apidés maintiennent constante leur vitesse optique de défilement du sol à (8 m/s) que divise (2 m), soit une vitesse optique de 4 rad/s (ou 230°/s) quelles que soient les variations du relief du terrain (figure 1). Si cette vitesse optique augmente, cela indique à l’insecte qu’il est en train de perdre de l’altitude, si cette vitesse optique diminue, cela indique à l’insecte qu’il est en train de gagner de l’altitude.
C’est donc en régulant cette information visuelle à un niveau constant que l’insecte opère son vol de croisière à une hauteur constante au-dessus du sol. L’abeille dispose donc uniquement d’un altimètre optique et non d’un altimètre barométrique comme les oiseaux, les poissons, ou bien les engins aériens construits par l’Homme.
L’étude publiée dans Biology Letters en 2022 (Serres et al., 2022) visait à pousser les connaissances sur le rôle des entrées visuelles provenant des ommatidies (lien) dorsales et ventrales alimentant le contrôle d’altitude des abeilles domestiques. Pour se faire, une équipe de chercheurs de l’Institut des Sciences du Mouvement – Étienne-Jules Marey UMR7287 – a imaginé manipuler cette vitesse optique de défilement du sol en l’annulant pour observer si les abeilles pouvaient encore voler dans cette invraisemblable configuration environnementale construite à l’aide de miroirs (figure 2).
L’abeille se fie-t-elle uniquement à la vitesse optique de défilement du sol pour contrôler son altitude ?

Aujourd’hui, un oui ferme nous apparaît comme irréfutable. Pour cela, les chercheurs se sont inspirés des expériences de Duchon & Warren (2002) conduites chez les humains, dans lesquelles une manipulation optique avait été construite par une paire de murs infinis (système de miroirs reflétant à l’infini un objet) afin de supprimer optiquement le sol, diminuant ainsi fortement les informations visuelles provenant du sol. Les chercheurs marseillais ont alors conçu une nouvelle configuration optique avec des miroirs au sol et/ou au plafond permettant de manipuler spécifiquement l’information de vitesse optique (figure 2), donc sans pour autant manipuler ou fausser d’autres informations optiques.
Une telle manipulation optique (figure 2), dans laquelle le sol semble avoir disparu, permet d’imiter certaines des conditions que connaissent les abeilles lorsqu’elles sont entraînées expérimentalement à voler au-dessus d’un plan d’eau naturel et permet ainsi étendre les connaissances de l’expérience fondatrice de l’entomologiste autrichien Herbert Heran (1920-1992) et de l’éthologiste allemand Martin Lindauer (1918-2008) conduite en 1963.
Cette expérience jamais reproduite depuis 60 ans, et parfois oubliée de la communauté des entomologistes, consistait à entraîner des abeilles à voler au-dessus d’une surface d’eau de 247 m de longueur. Lorsque cette surface était ondulée ou lorsqu’un pont flottant offrait un contraste visuel, les abeilles étaient capables de traverser le lac (figure 3). Par contre, les abeilles survolant une surface d’eau calme pendant leurs trajets aller volaient alors de plus en plus bas jusqu’à ce qu’elles entrent en collision avec la surface de l’eau puis se noyaient (Heran & Lindauer, 1963). Le tunnel de vol exploité dans l’étude publiée dans Biology Letters en 2022 (Serres et al., 2022) utilise une paire de miroirs placés au sol et au plafond qui se fond face, pour tenter de se rapprocher et de vérifier expérimentalement les observations comportementales en extérieur au-dessus d’une surface d’eau faites voilà 60 ans.

-
Matériels et méthodes
(a) Tunnel expérimental
Le tunnel de vol extérieur est de forme rectangulaire (220 cm de long, 71 cm de haut et 25 cm de large), le plafond et le plancher étant des miroirs qui peuvent être recouverts par des bâches texturées (figure 4). Le mur de gauche est constitué d’une planche alors que le mur de droite est constitué d’un filet anti-insecte. Un motif unique à rayures rouge et blanches, perpendiculaire à l’axe longitudinal du tunnel et donc orthogonalement à la direction du vol des abeilles, est disposé sur les quatre surfaces du tunnel (sol, plafond, paroi gauche, filet anti-insecte).
Ce motif n’est installé de manière permanente que sur les murs. Les miroirs du sol et du plafond peuvent être recouverts avec le même motif que le filet anti-insecte. Dans ce dernier cas, les rayures sont reproduites à l’aide d’un filtre à gélatine rouge (Lee Filters HT019). Le tunnel est fermé par des panneaux blancs à chaque extrémité. À l’une des extrémités, il y a une entrée circulaire (5 cm de diamètre) située à 11,5 cm du sol. À l’autre extrémité, une ouverture carrée (3,5 cm) placée à 11,5 cm au-dessus du sol permet aux abeilles d’accéder à la boîte. L’entrée du tunnel et la porte de la boîte de récompense s’ouvrent et se ferment manuellement par l’expérimentateur. Le tunnel de vol ne reçoit qu’un éclairage indirect (pas de lumière directe du soleil).

Cette vidéo peut être visionnée pour mieux comprendre l’organisation de notre dispositif expérimental :
(b) Motif périodique tapissant les parois du tunnel
Des bandes rouges de deux largeurs différentes (1 cm et 3 cm) forment un motif simple de 10 cm de large répété régulièrement. Les périodes angulaires des bandes rouges verticales varient de 5,7° à 53° (motif de 1-10 cm de large vu à une distance de 10 cm) et de 0,5° à 5,3° (motif de 1-10 cm de large vu à 1 m). Comme les abeilles ne possèdent pas de photorécepteurs sensibles au rouge (Srinivasan, 2011), elles perçoivent les bandes rouges comme des bandes grises. Entre les bandes rouges et blanches, le contraste de Michelson est de 0,47 mais de 0,25 sur le filet anti-insecte. Le contraste a été mesuré à l’aide d’une photodiode équipée d’un filtre passe-bande vert (Kodak Wratten n°61), dont le spectre de transmission correspond étroitement à la sensibilité spectrale des récepteurs verts de l’abeille.
-
Résultats
(a) Les abeilles qui suivent le sol ne s’appuient pas sur les informations visuelles dorsales
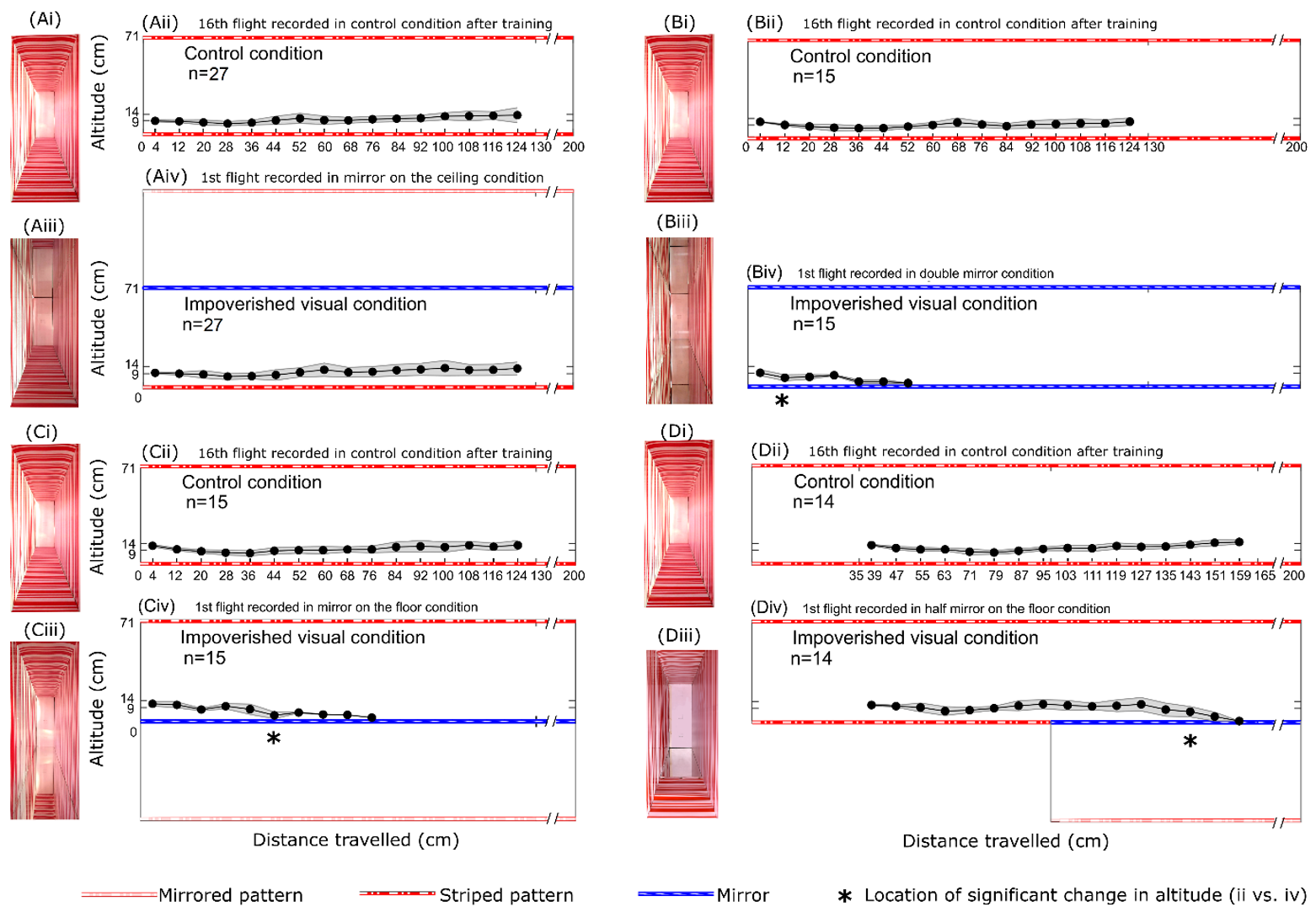
Dans l’expérience A (figure 5A), nous avons testé l’effet d’un appauvrissement visuel dans la partie dorsale de la vision des abeilles. Un groupe de 27 abeilles a été entrainé dans la condition de contrôle (figure 5Ai-ii). Le premier vol » en miroir sur le plafond plafond « , dans lequel le miroir du plafond était découvert, a été enregistré dans une condition visuelle appauvrie. La présence du miroir au plafond semblait doubler la hauteur du tunnel (142 cm) vers le haut (figure 5Aiii-iv). Nous n’avons alors observé aucun changement significatif dans le comportement de vol des abeilles domestiques.
(b) Sans aucune information visuelle ventrale et dorsale, les abeilles entrent en collision avec le miroir du sol.
Dans l’expérience B (figure 5B), nous avons testé l’effet d’un appauvrissement visuel dans les parties dorsale et ventrale du champ visuel des abeille. Un groupe de 15 abeilles a été entraîné dans la condition de contrôle (figure 5Bi-ii). Le premier vol, condition « en double miroir » dans laquelle les deux miroirs étaient découverts, a été enregistré dans une condition visuelle appauvrie. La présence de miroirs au plafond et au sol a créé une manipulation optique dans laquelle une paire de murs infinis est apparue. Le plancher ou le plafond n’était plus disponible (figure 5Biii-iv). Nous avons observé des changements significatifs dans le comportement de vol des abeilles à partir de x = 8 cm jusqu’à ce que chacune des abeilles entre en collision avec le miroir du sol (figure 5Biv).
(c) Les informations visuelles dorsales n’aident pas à voler plus loin au-dessus du miroir du sol.
Dans l’expérience C (figure 5C), nous avons testé l’effet d’un appauvrissement visuel dans la partie ventrale du champ visuel des abeilles. Un groupe de 15 abeilles a été entraîné dans la condition contrôle (figure 5Ci-ii). Le premier vol, condition « miroir au sol » dans laquelle le miroir au sol était découvert, a été enregistré dans une condition visuelle appauvrie. La présence du miroir sur le sol semble doubler la hauteur du tunnel (142 cm) vers le bas (figure 5Ciii-iv), créant une sorte de « fossé ventral » de 71 cm de profondeur. Nous avons observé des changements significatifs dans le comportement de vol des abeilles à partir de x = 40 cm jusqu’à ce que chacune des abeilles entre en collision avec le miroir du sol (figure 5Civ). Les abeilles peuvent être visuellement attirées par le sol virtuel situé 71 cm plus bas, mais elles sont ensuite entrées en collision avec le miroir.
Cette vidéo peut être visionnée pour observer qualitativement comment se sont comportées les abeilles en conditions contrôlées :
Cette vidéo peut être visionnée pour observer qualitativement comment se sont comportées les abeilles au-dessus d’un miroir :
(d) Recouvrir la première moitié du miroir de sol n’aide pas les abeilles à voler plus loin.
Dans l’expérience D (figure 5D), nous avons testé l’effet d’un renforcement visuel dans la partie ventrale du champ visuel des abeilles en recouvrant la première moitié du miroir au sol avec le même motif de rayures rouges et blanches que celui utilisé précédemment. Un groupe de 14 abeilles a été entraîné dans la condition contrôle (figure 5Di-ii). La première condition de vol « dans la moitié du miroir au sol » dans laquelle la seconde moitié du miroir au sol a été laissée découverte double virtuellement la hauteur du tunnel (142 cm) vers le bas (figure 5Diii-iv) créant ainsi une sorte de « fossé ventral » de 71 cm de profondeur comme dans l’expérience C (figure 5C). Nous avons observé des changements significatifs dans le comportement de vol des abeilles à partir de x = 139 cm jusqu’à ce que chacune des abeilles entre en collision avec le miroir (figure 5Div). Cette texture supplémentaire sur le sol n’aide donc pas les abeilles à voler plus loin au-dessus du miroir. La distance parcourue au-dessus du miroir (39 cm) était similaire à celle observée dans l’expérience C (figure 5Civ).
-
Discussion et conclusion
Dans l’expérience A (figure 5Aiv), les abeilles semblent suivre le sol malgré le « fossé dorsal » virtuel, car elles ont peut-être appris à suivre le sol et à se fier aux informations visuelles ventrales pour réguler leur vol, ce qui concorde avec les résultats de Portelli et al. (2017) où les abeilles ne sont pas influencées par la hauteur du tunnel dès lors qu’elles suivent le sol. L’absence de différence entre le dernier vol d’entraînement et les données de l’essai de l’expérience A suggère que le changement de hauteur du tunnel vers le haut n’a peut-être tout simplement pas été perçu par les abeilles.
Inversement, chaque manipulation expérimentale qui affecte la partie ventrale du flux optique, qu’il s’agisse d’une privation totale du flux ventral dans l’expérience B (figure 5Biv) ou d’une réduction de la partie ventrale du flux (figure 5Civ et figure 5Div), a donné lieu à une perte d’altitude jusqu’à ce que l’abeille entre en collision avec le miroir du sol. Il est intéressant de noter que notre condition de double miroir nous a permis de nous rapprocher des conditions d’un vol en plein ciel au-dessus d’une surface d’eau calme, comme le modèle utilisé par Heran et Lindauer (1963). Nos résultats concordent avec les leurs dans la mesure où les abeilles perdent de l’altitude en l’absence de flux optique ventral.
Les résultats sont sans appel : sans cette vitesse de défilement optique du sol, les abeilles tombent ou rebondissent sur le miroir, certainement à la recherche du sol.
Toute l’originalité de ces travaux réside dans la conception de configurations optiques contrôlées et manipulables se rapprochant au mieux des conditions naturelles de vol des abeilles, sans pour autant avoir besoin d’un lac ou de sacrifier des abeilles pour étudier le contrôle d’altitude.
Les chercheurs marseillais devaient être certain de pouvoir mettre en défaut la régulation d’altitude des abeilles pour pouvoir ensuite leur proposer d’autres informations optiques, puis étudier leur combinaison pour décrire le contrôle d’altitude (Berger Dauxère et al., 2021). Notre dispositif expérimental pourrait être également utilisé pour évaluer la perception de la vitesse optique de défilement du sol chez d’autres insectes volants et ainsi en observer, ou non, le caractère universel, ou non.
Vous pouvez retrouver une présentation de ces résultats en anglais sur ma chaîne Youtube :
Auteur
 Julien est Maître de Conférences au sein de l’Institut des Sciences du Mouvement – Etienne-Jules Marey (CNRS / Aix Marseille Université, UMR7287, https://ism.univ-amu.fr/). Il s’intéresse au système de navigation des insectes non seulement pour mieux comprendre comment les insectes perçoivent leur environnement mais aussi pour développer de nouvelles techniques de localisation biomimétique à des fins applicatives en robotique.
Julien est Maître de Conférences au sein de l’Institut des Sciences du Mouvement – Etienne-Jules Marey (CNRS / Aix Marseille Université, UMR7287, https://ism.univ-amu.fr/). Il s’intéresse au système de navigation des insectes non seulement pour mieux comprendre comment les insectes perçoivent leur environnement mais aussi pour développer de nouvelles techniques de localisation biomimétique à des fins applicatives en robotique.
Ses liens professionnels :
– https://www.linkedin.com/in/julien-serres-945a27115/
– https://www.youtube.com/channel/UC5fXQaHieW6yGj4mUKuB_7Q/videos
– Site professionnel : https://serres-lab.com/equipe/)
Bibliographie
- Berger Dauxère, A., Serres, J. R., & Montagne, G. (2021) : Ecological entomology: how is Gibson’s framework useful? Insects, 12(12), 1075 (lien)
- Duchon, A. P. & Warren Jr, W. H. (2002) : A visual equalization strategy for locomotor control: of honeybees, robots, and humans. Psychological Science, 13(3), 272-278 (lien)
- Lecoeur, J., Dacke, M., Floreano, D. & Baird, E. (2019) : The role of optic flow pooling in insect flight control in cluttered environments. Scientific reports, 9(1), 1-13 (lien)
- Heran, H. & Lindauer, M. (1963) : Windkompensation und seitenwindkorrektur der bienen beim flug über wasser. Zeitschrift für vergleichende Physiologie, 47(1), 39-55 (lien)
- Portelli, G., Ruffier, F. & Franceschini, N. (2010) : Honeybees change their height to restore their optic flow. Journal of Comparative Physiology A, 196(4), 307-313 (lien)
- Portelli, G., Serres, J. R. & Ruffier, F. (2017) : Altitude control in honeybees: joint vision-based learning and guidance. Scientific reports, 7(1), 1-10 (lien)
- Riley, J. R. & Osborne, J. L. (2001) : Flight trajectories of foraging insects: observations using harmonic radar. Insect Movement: Mechanisms and Consequences (ed. I. P. Woiwood, D. R. Reynolds and C. D. Thomas), pp.129-158. Wallingford: CABI Publishing
- Srinivasan, M. V. (2011) : Honeybees as a model for the study of visually guided flight, navigation, and biologically inspired robotics. Physiological reviews, 91(2), 413-460 (lien)
- Serres, J. R., Morice, A. H., Blary, C., Miot, R., Montagne, G. & Ruffier, F. (2022) : Floor and ceiling mirror configurations to study altitude control in honeybees. Biology Letters, 18(3), 20210534 (lien)
- Von Frisch, K. (2013). The dance language and orientation of bees. In The dance language and orientation of bees. Cambridge, MA: Harvard University Press