Les effets de la dispersion et de la migration sur la morphologie alaire du Monarque
Par Benoît GILLES
Pour les animaux, réaliser des migrations sur de longues distances confère de nombreux avantages comme la possibilité d’exploiter des ressources alimentaires saisonnières et éphémères. C’est pourquoi ces comportements sont apparus et ont évolué à de multiples reprises au cours de l’évolution.
La sélection a apporté aux espèces migratrices une combinaison de traits morphologiques, physiologiques et des capacités d’orientation leurs permettant de se déplacer sur de longues distances.
A l’inverse, la perte du comportement migratoire est également un processus courant, à la fois à l’échelle de l’évolution mais aussi de manière contemporaine. Lorsque des espèces passent du statut de migratrices à celui de sédentaires, les traits phénotypiques qui étaient auparavant importants pour les déplacements ne sont plus sélectionnés, voire contre-sélectionnés, car physiologiquement coûteux à maintenir, au détriment d’autres caractères plus favorables à la sédentarisation.

Pour étudier ces changements et comprendre comment la sélection façonne les gènes et les traits associés, il est possible de s’intéresser à des mécanismes s’étalant sur plusieurs millions d’années à l’aide des fossiles, ou d’avoir recours à des espèces dont les changements se sont déroulés sur un laps de temps court.
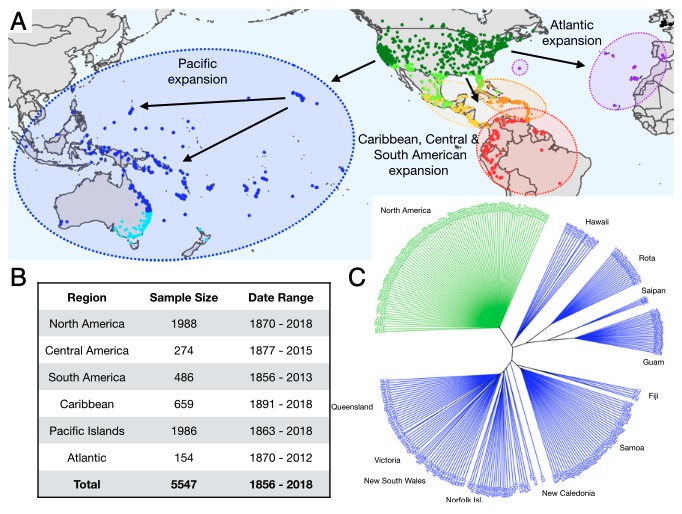
Le Monarque (Danaus plexippus) est un papillon connu pour ses migrations annuelles multigénérationnelles en Amérique du Nord, centrale et du sud, dans les Caraïbes, où certains individus parcourent près de 3000 km au cours de leur vie (figure 1). Il se rencontre aujourd’hui dans certaines régions du Pacifique (îles du Pacifique, Indonésie, Australie) et de l’Atlantique suite à de multiples événements d’introduction d’origine anthropique au cours des 180 dernières années (figure 2 A ci-dessous).
Les populations d’Amérique centrale et du sud se seraient, quant à elles, établies il y a une dizaine de milliers d’années, formant aujourd’hui des populations génétiquement distinctes.
Dans presque toutes les régions où le Monarque s’est établi, les populations ont perdu leur faculté à migrer en raison de la disponibilité de leur plante hôte (asclépiades) tout au long de l’année. Ainsi, à chaque événement majeur d’expansion hors de l’Amérique du Nord correspond une perte de migration indépendante offrant des preuves d’une évolution génomique convergente associée à cette transition.
Un phénotype favorable à l’expansion
En raison de sa coloration, le Monarque a souvent fait l’objet de collectes par des entomologistes à travers la planète, fournissant de ce fait d’excellents enregistrements des phénotypes (variations de la coloration et de la morphologie des ailes) en fonction de la géographie et de la date de colonisation de la nouvelle aire (autre modèle d’études : les papillons Heliconius).
Des études ont, par le passé, mis en évidence un taux de dispersion élevé le long du front d’expansion chez de nombreuses espèces. Ce phénomène a été particulièrement bien documenté pour le crapaud buffle (Rhinella marina) introduit en Australie en 1935 pour lutter contre des Coléoptères s’attaquant aux cannes à sucre. Les crapauds se trouvant en périphérie de l’aire d’expansion avaient des pattes plus longues et se déplaçaient sur de plus longues distances que ceux situés au centre de l’aire d’expansion. Ce phénomène a été mis en évidence également pour des plantes ou des oiseaux océaniques dont les individus fondateurs présentent des traits favorables à la dispersion.
Des preuves génétiques attestent de la dispersion progressive des Monarques dans le Pacifique et l’Atlantique. L’Américain Micah G. Freedman et son équipe ont souhaité mettre en évidence une relation entre la propension à la dispersion et l’évolution des phénotypes des papillons au cours du temps (figure 2). Il a émis l’hypothèse que la sélection aurait entrainé une réduction de la capacité de dispersion résultant : 1) soit d’une sélection directionnelle opérant sur un ou plusieurs traits potentiellement corrélés pour favoriser un nouvel optimum non migratoire ; 2) soit d’un relâchement de la sélection directionnelle ou stabilisatrice précédemment associée à un optimum de trait migratoire (voir encadré ci-dessous).
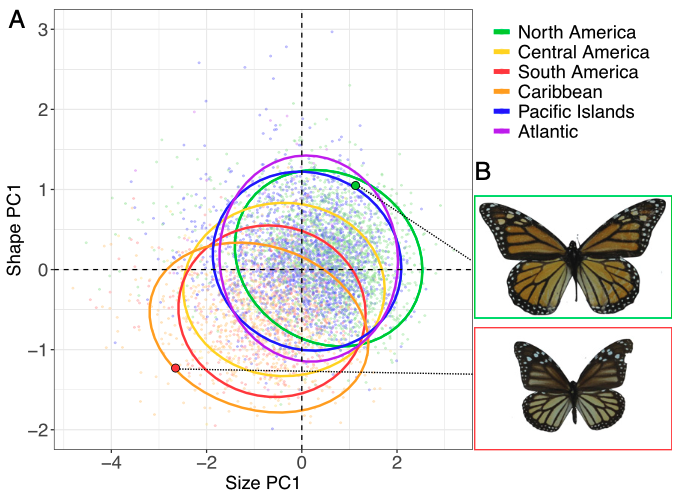
Des études antérieures ont établi que la taille des ailes antérieures des populations de Monarques non migrateurs était plus petite et qu’elles étaient moins allongées. Cependant, ces études n’ont pas pris en compte la récente perte de la capacité migratoire dans le Pacifique et l’Atlantique, ni le rythme auquel cette perte a été opérée.
Pour considérer ces paramètres, l’équipe a entrepris d’analyser la morphologie des ailes de 6 000 spécimens de collections historiques datés à partir de 1856.
Phénotype et capacité à migrer
Les résultats de l’étude indiquent que les premiers spécimens arrivés dans le Pacifique et l’Atlantique avaient des ailes antérieures significativement plus grandes et allongées que ceux des populations ancestrales nord-américaines (figure 3 ci-dessous). La même observation a été réalisée sur les spécimens migrant à la limite nord de leur aire de répartition en Amérique du Nord qui possèdent également des ailes antérieures de plus grande taille et plus allongées que ceux situés au centre de leur aire de répartition.
Des spécimens sont parfois observés et collectés au Royaume-Uni après le passage de tempêtes : provenant d’Amérique du Nord bien que ne pouvant pas s’établir par absence de leur plante-hôte, cela démontre la capacité naturelle de dispersion de ces insectes !
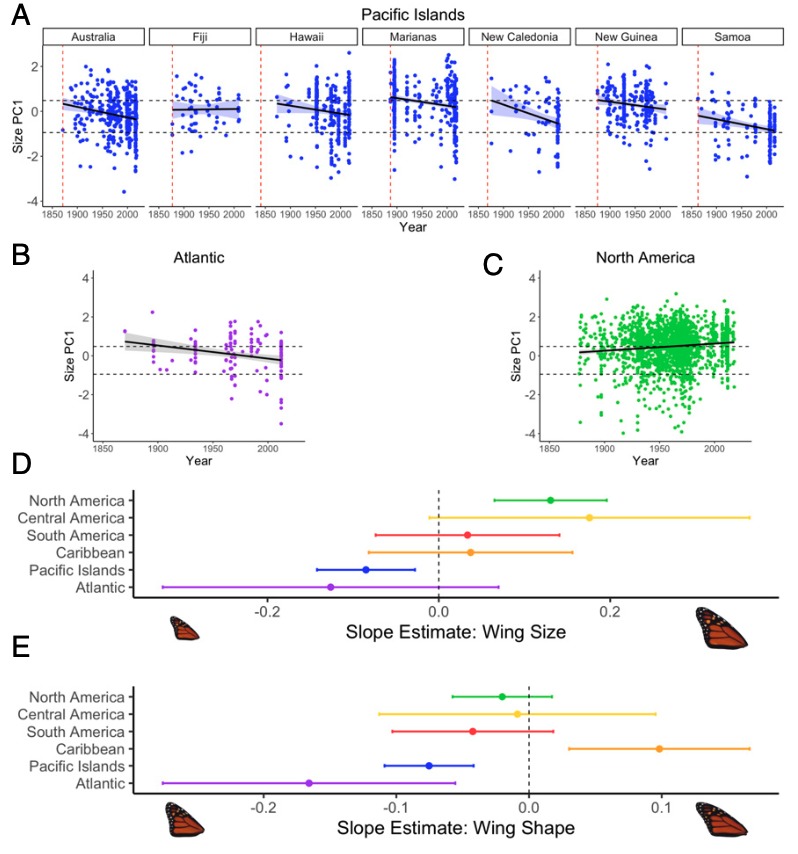
L’étude a également mis en évidence chez les populations du Pacifique et de l’Atlantique non migratrices une réduction au cours du temps de la taille des ailes antérieures et de leur allongement par rapport aux populations fondatrices (figure 4). En revanche, la taille des ailes des Monarques migrateurs nord-américains a, quant à elle, augmenté depuis 1870.
Par contre, les populations d’Amérique centrale, du Sud et des Caraïbes, certainement non migratrices depuis plusieurs milliers d’années, ne présentent pas d’évolution de la morphologie de leurs ailes.
De manière surprenante, la taille des ailes des spécimens non migrateurs d’Amérique centrale a augmenté, certainement en raison de flux génétiques réguliers entre les populations d’Amérique du nord et celles d’Amérique centrale, un scénario soutenu par une faible différenciation génétique entre ces populations.
Types de sélection
Ce type de sélection favorise les phénotypes extrêmes et ce, dans une direction ou une autre, pour évoluer vers un optimum. Il est souvent rencontré lorsqu’une population subit des changements extrinsèques comme des changements environnementaux ou lorsqu’une population émigre dans un nouvel habitat non identique à l’original.
Ce type de sélection se produit lorsque les phénotypes extrêmes ont un avantage sur les phénotypes intermédiaires, la proportion de ces derniers ayant tendance à diminuer. Cette sélection mène potentiellement à une spéciation sympatrique (émergence de deux espèces à partir d’un ancêtre commun sans isolement géographique). Elle peut être de deux types :
Contrairement aux deux autres types de spéciation qui favorisent les phénotypes extrêmes, celle-ci élimine ces derniers pour favoriser les intermédiaires, un processus ayant pour effet de diminuer la variance du caractère entre individus et de maintenir le statu quo relatif à un phénotype spécifique. |
Des modifications morphologiques d’origine génétique
Des expériences menées en condition d’élevage sur des populations de Monarques non migrateurs provenant d’Hawaï, de l’île de Guam, d’Australie et de Porto Rico ont montré que ces papillons avaient des ailes antérieures de plus petite taille que ceux des populations migratrices d’Amérique du nord (figure 4). Ces observations semblent indiquer que la variabilité de la taille des ailes résulterait plus de facteurs génétiques que de la plasticité phénotypique associée aux différences environnementales des régions géographiques d’implantation.
De plus, aucune différence morphologique au niveau de la masse corporelle, de la charge alaire (proportion des ailes par rapport à la taille du corps) et de l’allocation de masse des tissus thoraciques (investissement et coûts énergétiques en lien avec l’utilisation et l’entretien des muscles du thorax) n’a été constatée entre les populations migratrices et non migratrices.
Il semble également que les populations de Monarque d’Amérique de l’est et de l’ouest constituent une unique population génétiquement indifférenciée sur tout le continent nord-américain.
Des analyses suggèrent que les niveaux de divergence observés entre les phénotypes des populations d’Amérique du nord et du Pacifique peuvent être expliqués principalement par la dérive génétique (perte de la diversité génétique associée à une population en raison d’un faible nombre d’individus passant d’une génération à l’autre), hypothèse plus cohérente que celle d’une sélection survenue à la suite d’un événement d’établissement (installation d’individus dans une nouvelle région).
Conclusion
A l’aide de données provenant de spécimens en collection et d’expériences en conditions d’élevage, l’équipe de Micah G. Freedman a démontré que la dispersion des Monarques dans de nouvelles zones géographiques a été effectuée par des individus possédant de grandes ailes, puis, une fois établies, que la taille des ailes s’est réduite au cours d’environ 1 000 générations.
Ces résultats intéressants offrent 1) une preuve d’un lien direct entre la capacité d’expansion et la perte de la faculté à migrer ; 2) une chronologie du changement morphologique dans le temps et du mode de sélection sous-jacent.
Il est en effet complexe de faire la distinction entre la sélection directionnelle vers un optimum phénotypique de type non migratoire et la sélection de traits maintenus par la migration : la perte des yeux chez les poissons cavernicoles, la réduction de la colonne vertébrale chez les épinoches à trois épines, ou encore la perte du vol chez les oiseaux insulaires par exemple.
Les ailes du Monarque interviennent dans de nombreux comportements en plus de la migration : recherche de nourriture, évitement de prédateurs, poursuite de partenaires sexuels par exemple. L’absence de données sur la structuration génétique (gènes et loci) associée à la morphologie de l’aile, il est actuellement difficile de déterminer directement si les populations non migratrices résulteraient d’un processus de sélection divergente (perte de fonction) ou de sélection directionnelle (perte de diversité génétique).
Les données phénotypiques obtenues durant l’étude fournissent des preuves confirmant la première hypothèse. Le rythme de l’évolution morphologique des ailes a été graduel. Arrivés en 1871, les Monarques australiens possèdent une surface alaire 7,3% plus petite que celle de leurs ancêtres nord-américains (819 mm2 contre 884 mm2), soit une réduction de moins de 0,5 mm2 par an, ou de moins de 0,1 mm2 par génération.
Ces résultats n’excluent pas la possibilité que le caractère « ailes antérieures larges » soit activement sélectionné dans les populations non migratrices. Des recherches complémentaires sont nécessaires pour comprendre comment des facteurs autres que la migration à longue distance façonnent la morphologie des ailes des Monarques.
Enfin, cette étude met en évidence l’importance des collections pour tester des hypothèses évolutives.
Les collections ont fourni non seulement les spécimens utilisés pour les mesures mais aussi permis de déduire l’histoire expansive du Monarque dans l’Atlantique et dans le Pacifique. Comme l’expansion de l’aire de répartition de nombreuses espèces associée au changement climatique et aux activités anthropiques devient courante, la valeur des collections de spécimens biologiques ne fera qu’augmenter.
Bibliographie
- Freedman M.G. ; Dingle H. ; Strauss S.S. & Ramirez S.R. (2020) : Two centuries of monarch butterfly collections reveal contrasting effects of range expansion and migration loss on wing traits. Proceedings of the National Academy of Sciences, 1-7 (lien)