Électricité statique et insectes pollinisateurs
Par Benoît GILLES
Le monde des insectes n’a pas fini de nous surprendre. En plus de capacités physiologiques, écologiques et comportementales exceptionnelles leur permettant de s’adapter à tout type de milieu et d’habitat, les insectes se révèlent avoir également la capacité de détecter et d’utiliser les champs d’électricité statique des fleurs pour identifier leur potentiel de récompense (nectar ou pollen).
Les premières observations de ces charges d’électricité statique chez les insectes remontent à 60 ans. Toutefois, c’est en 2013 que l’entomologiste anglais de l’Université de Bristol (Grande-Bretange) Clarke, et ses collègues (Clarke et al., 2013), à l’aide d’un appareillage de mesure adapté, ont réussi à les évaluer. Il s’avère qu’avec 32 pC (soit x10-8 coulomb – picocoulomb) (un coulomb correspondant à la quantité de charge électrique transférée par un courant d’un ampère en une seconde), ce sont les Bourdons terrestres (Bombus terrestris) les champions en termes de production de charges électrostatiques.
Insectes, fleurs et électricité statique
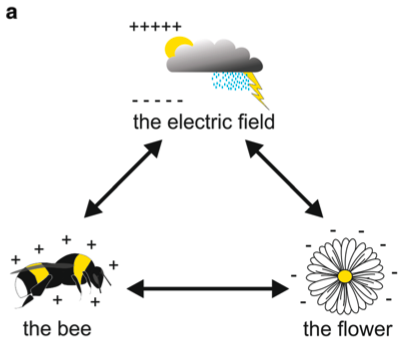
Les charges d’électricité statique de l’insecte résultent des frottements des ailes et des autres structures de son corps avec l’air de la même manière que des cheveux humains le font avec un peigne en plastique. Ce phénomène mal compris, appelé « effet triboélectrique », se produit lorsque deux matériaux entrent en contact l’un avec autre. Des électrons sont alors transférés entre les deux surfaces mais lors de la séparation, une partie des électrons ne retrouvent pas leur position initiale à la surface des matériaux. Cette différence provoque un déséquilibre résiduel : une charge électrostatique positive apparaît alors.
Il semblerait que les Bourdons soient fortement chargés en électricité statique en raison de 1) leur grande taille qui offre une large surface de cuticule avec l’air, notamment au niveau du thorax et des ailes ; 2) leur forte pilosité jouant un rôle d’isolant thermique qui leur permet de voler à des températures ambiantes plutôt basses (Heinrich, 1976). Les Bourdons femelles sont en effet visibles dès la fin de l’hiver et au début du printemps à la recherche de nourriture et d’un site où installer leur nid.
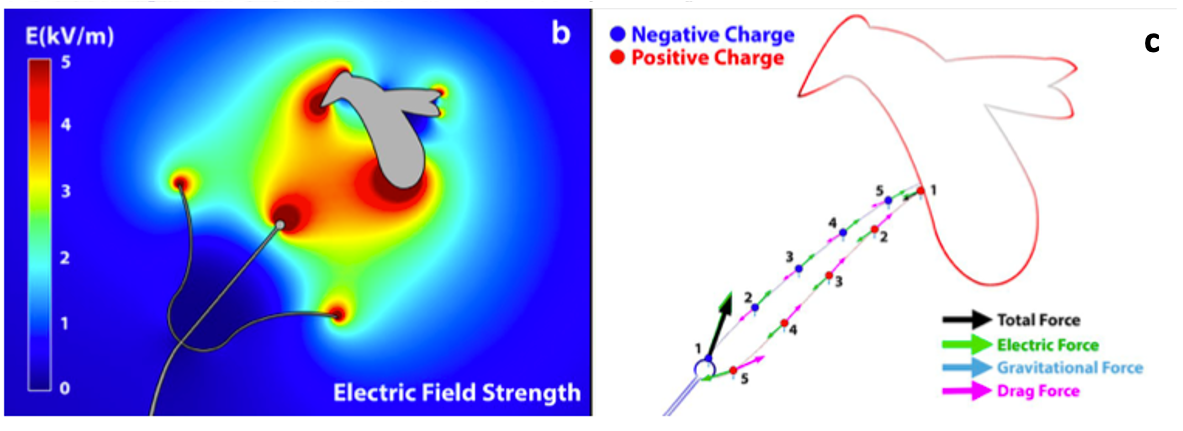
Ainsi, durant le vol un Bourdon génère une charge positive nette par le biais d’interactions frictionnelles avec des particules de l’air et de parties de son corps entre elles comme les pattes ou les ailes avec le thorax (figure 1).
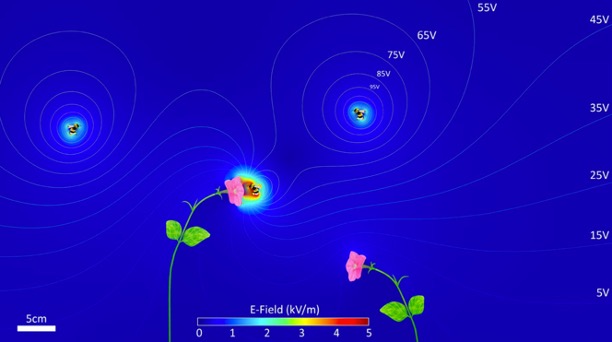
Les plantes sont également porteuses de charges électrostatiques de l’ordre de 100 Vm-1, avec une intensité de champ atteignant des valeurs beaucoup plus élevées (jusqu’à 3 kVm-1) près des structures « pointues » telles que les pétales et les organes reproducteurs des fleurs (Bowker et al., 2007). Plus la fleur éloignée du sol, plus son potentiel d’électrique statique sera élevé par rapport à l’air environnant (England et al., 2021) (figure 1).
Système de marquage électrostatique
Plantes à fleurs et insectes pollinisateurs co-évoluent depuis l’apparition des fleurs il y a environ 275 millions d’années (Solomo et al., 2017). Cette relation mutualiste repose sur le fait que les plantes à fleurs sont pollinisées par des insectes qui assurent le transfert des grains de pollen (gamètes mâles) d’une fleur à une autre en échange de récompenses nutritionnelles fournies par la plante sous forme de nectar et de pollen (lien).
Le transfert des grains de pollen de la fleur à l’insecte, et inversement, se fait par simple contact mécanique. Pour faciliter leur transport d’une fleur à l’autre, les grains possèdent une surface adhésive ou une surface composée de « crochets ».
En 1982, l’équipe de Corbet (Corbet et al., 1982) a pour la première fois mis en évidence que des charges électrostatiques intervenaient dans le processus de transfert des grains de pollen de la fleur à l’insecte et vice-versa sans recours à un contact physique ou mécanique. En effet, il a été mesuré qu’une différence de potentiel électrique pouvait propulser des grains de pollen dans un espace d’air de plusieurs mm entre une fleur de colza et un bourdon immobilisé, la distance du transfert étant proportionnelle au carré de la différence de tension (figure 3).
A l’aide de modèles mathématiques réalistes (Vaknin et al., 2000 ; Clarke et al., 2017) et d’expérimentations consistant à soupoudrer des fleurs avec une poudre colorée et chargée électrostatiquement (Clark et al, 2017), il a été mis en évidence que le champ électrique à la surface des fleurs n’est pas uniforme mais présente des intensités plus élevées (jusqu’à 5 kVm-1) à des endroits spécifiques comme l’intérieur et les extrémités des pétales, et au niveau des structures reproductrices (étamines et pistils) (figures 2 & 3).

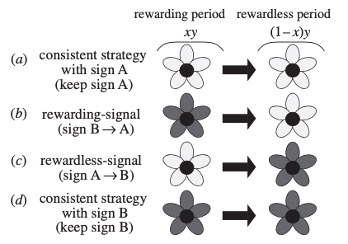
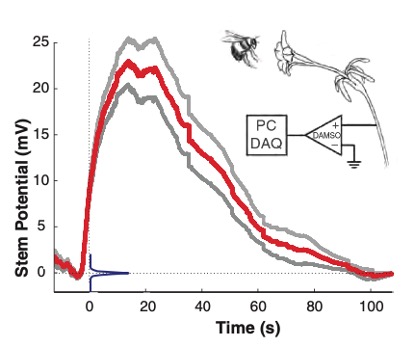
Autre observation, lorsqu’un Bourdon terrestre (Bombus terrestris) visite une fleur de Pétunia, le contact provoque une polarisation positive de la fleur par rapport à son potentiel normal. Ce changement de potentiel augmente rapidement, atteint son pic 20 secondes après la visite et revient à une valeur initiale au bout de 100 secondes environ (figure 8) (Clarke et al., 2013).
Ces observations suggèrent l’existence d’un mécanisme évolutif faisant pression sur la forme des fleurs pour favoriser le transfert des grains de pollen sur les insectes par voie électrostatique (Arnbruster, 2001 ; Vaknin et al., 2001).
La maitrise de ces processus pourrait être avantageux, en termes évolutifs, pour la plante en améliorant le transfert des grains de pollen d’une fleur à autre plante via les insectes (zoochorie) et l’être aussi pour les insectes pollinisateurs en facilitant la collecte de ressources alimentaires.
Détection et interprétation des marqueurs électrostatiques par les insectes pollinisateurs
Pour un insecte pollinisateur, avoir la capacité de détecter et d’interpréter les variations de charges électrostatiques à la surface des fleurs peut être particulièrement utile et avantageux dès lors que cela lui permette d’évaluer si la fleur a été précédemment visitée par un autre individu, et la quantité de nourriture disponibles (pollen et nectar).
Depuis les années 1960, bien qu’il soit admis qu’un grand nombre de taxons d’insectes soient sensibles à l’électricité statique, aucune preuve ne permettait de démontrer que les insectes avaient la capacité d’attribuer une signification à la charge électrique émise par la fleur.
Les travaux de l’équipe de Clarke (2013) ont apporté les premières preuves d’une sélection des fleurs en fonction des charges électrostatiques chez les Bourdons terrestres (Bombus terrestris).
Les expérimentations ont consisté à 1) positionner des fleurs artificielles en résine époxy pour lesquelles les charges d’électricité statiques et les types de « récompenses » alimentaires pouvaient être modulés ; 2) observer les comportements des insectes ; 3) identifier les paramètres utilisés par les Bourdons pour réaliser leur choix (figure 4).
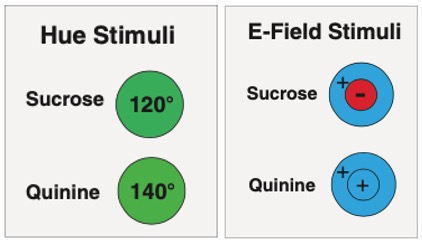
Face à des fleurs présentant une tension continue de +30 V et une récompense sucrée, et à des fleurs au goût amer (quinine) non chargées (potentiel zéro), les Bourdons ont été, dans un premier temps, incapables d’identifier la source de nourriture (sucres) (50% des visites vers les fleurs +30 V et 50% aux fleurs 0 V). Cependant, les insectes ont rapidement appris à visiter les fleurs chargées électriquement contenant du sucre : après 50 essais, 80% de leurs visites concernaient des fleurs portant le potentiel électrique +30 V.
La suppression de la charge électrique a rendu les Bourdons incapables de distinguer les fleurs sucrées des fleurs amères, et cela même avec une tension réduite à seulement +10 V (figure 2b).
Ces expérimentations démontrent que les Bourdons identifient les sources de nourriture grâce à leur capacité de détection de l’état électrique des fleurs, les charges devant toutefois dépasser une certaine valeur (effet de seuil).
Contrairement aux fleurs naturelles, les fleurs artificielles ne présentaient pas des patterns de charges uniformes. Pour éliminer ce biais, Clarke et son équipe ont construit des fleurs artificielles portant soit 1) une tension uniforme de +20 V et contenant une « récompense » de quinine aversive, 2) un schéma électrique en forme de cible (un anneau à +20 V entourant un centre à -10 V) et contenant une « récompense » sucrée (figure 4).
Comme précédemment, les Bourdons ont rapidement appris à différencier les deux modèles de fleurs. La suppression du signal électrique a une nouvelle fois provoquée l’incapacité des Bourdons à distinguer les deux types de fleurs.
Pour aller plus loin, cette équipe a ajouté un paramètre : la couleur. Elle a proposé des fleurs d’intensité de couleur différente (nuance de verts). Encore une fois, les Bourdons ont rapidement appris à distinguer et à préférer les fleurs artificielles contenant une récompense sucrée des fleurs artificielles sans récompense selon les intensités de couleur. Le choix était d’autant plus rapide lorsque les fleurs portaient une charge électrique (fig. 2d).
Ces recherches ont permis de mettre en évidence que les Bourdons possèdent la capacité d’intégrer et de coupler des informations à la fois sensorielles dérivées des champs électriques floraux et des informations visuelles reposant sur la couleur des fleurs.
Cela suggère que l’électro-réception est une partie inhérente du répertoire comportemental de pollinisation des Bourdons (figure 6).
Ces travaux amènent également à conclure que les bourdons utilisent une double source d’informations (électrique et visuelle) pour décider quelle fleur butiner. Les fleurs, de leur côté, envoient des informations aux pollinisateurs sur leurs valeurs nutritives.
Comment les insectes détectent-ils la présence de champs électriques statiques ?
Les signaux électrostatiques sont détectés et codés chez les bourdons par les poils mécanosensoriels thoraciques (figure 5) (Sutton et al., 2016) (lien). En effet, la présence d’un champ d’électricité statique provoque un déplacement latéral de l’air qui fait bouger les poils thoraciques. Chaque poil stimule alors le neurone situé à sa base ce qui déclenche des potentiels d’action qui gagnent le système nerveux central (lien). Le mouvement du poil et le potentiel d’action sont d’autant plus importants que la charge électrique du poil est élevée.
Des études comparatives entre les poils mécanosensoriels de B. terrestris et ceux d’autres espèces d’insectes ne montrent pas de particularité spécifique ni de différence mécanique ou autre, par rapport aux autres espèces d’insectes.
La capacité à détecter les champs électriques statiques pourrait être assez générale chez les insectes (Palmer et al., 2021).
En 2022, des expérimentations semblables à celles menées par l’équipe de Clarke sur B. terrestris ont également démontré la capacité à détecter et à interpréter les signaux des champs électriques chez deux espèces de Syrphes : Cheilosia albipila et Eristalis tenax, (Khan et al. 2021).

Avantages et coûts du marquage des fleurs par les pollinisateurs
Malheureusement, peu d’éléments concrets permettent d’affirmer avec certitude que les marqueurs électrostatiques présents à la surface des fleurs soient réellement utilisés dans la nature par les pollinisateurs pour guider leur choix de fleur lors de la recherche de nourriture.
Les questions qui se posent sont les suivantes :
- Quels avantages les insectes tirent-ils d’un message électrique ?
Tout d’abord, un grand nombre d’espèces pollinisatrices comme les abeilles mellifères (Apis mellifera), certaines espèces d’abeilles « solitaires » et de bourdons ont recours à des marqueurs chimiques pour signaler leur visite dans une fleur. Un système reposant sur l’électricité statique semble donc pouvoir évoluer de la même manière (Giurfa et al., 1992 ; Yokoi et al., 2009).
Pouvoir détecter si la fleur a déjà été visitée offre un avantage en termes de gain de temps et d’énergie : en évitant de visiter une fleur dont les ressources nutritives ont déjà été collectées, l’insecte améliore la rentabilité de l’activité de recherche de nourriture.
Premier cas de figure : une abeille arrive sur une parcelle de fleurs avec une trop grande proportion de fleurs récemment visitées, elle pourrait décider d’abandonner le site et de s’envoler vers une nouvelle parcelle.
Second cas de figure : le système permettrait à une abeille de s’assurer de ne pas perdre de temps en revisitant plusieurs fois les mêmes fleurs.
Cependant, pour être le plus efficient possible, un tel système de marquage doit être accessible et compris par une majorité des pollinisateurs.
Bourdon en train de butiner de fleur en fleur
- Pourquoi serait-il avantageux d’avoir deux systèmes ?
Il est difficile de savoir lequel des deux systèmes est apparu en premier.
Cependant, spontanément, le contact entre un insecte et les tissus de la fleur engendre une dépolarisation en raison des charges électrostatiques échangées.
Les mécanismes sensoriels et neuronaux préexistants chez les insectes pour appréhender leur environnement auraient pu être réutilisés pour permettre la détection et l’interprétation des signaux floraux. De plus, le système électrostatique de marquage des fleurs, probablement plus simple que le marquage olfactif puisqu’il ne nécessite pas la synthèse de molécules chimiques, certains scientifiques comme Stuart Reynolds (Département de Biologie et de Biochimie, Université de Bath) suggèrent que le marquage électrostatique des fleurs serait antérieur au marquage olfactif.
Une des possibilités est que les pollinisateurs puissent apprendre plus rapidement des caractéristiques communes à toutes les fleurs contrairement aux messages chimiques plus spécifiques de l’espèce végétale. Il a été démontré que les abeilles apprenaient plus rapidement et de manière plus efficace lorsque plusieurs modalités sensorielles étaient sollicitées (Leonard et al., 2011).
De plus, comme le suggère Stuart Reynolds, le signal électrique offre l’avantage d’être plus rapide que le signal chimique, tant dans son activation que sa disparition. En effet, le dépôt d’une charge électrique statique sur la fleur par l’insecte est un processus instantané survenant immédiatement avec le contact pour s’annuler quelques secondes plus tard.
Les différences à l’échelle temporelle des informations véhiculées par les deux systèmes de signalisation des récompenses suggèrent l’existence de types de message spécifiques, mais cela reste à confirmer.
Enfin, les signaux électrostatiques sont, contrairement aux messages chimiques, identifiables par tous les pollinisateurs car ils ne nécessitent pas de récepteurs spécifiques, bien qu’il puisse y avoir des différences entre les grandes et les petites espèces.
Cela signifie que la fiabilité des informations contenues dans un signal électrostatique de marquage floral peut être supérieure à celle d’un signal olfactif.
- Et la plante dans tout ça ?
Pour la plante, communiquer sur les visites des pollinisateurs et renseigner sur les quantités disponibles en ressources alimentaires ne semble pas forcément intéressant. En effet, pour optimiser les chances d’être fécondées, les fleurs doivent attirer le maximum de pollinisateurs. Offrir une récompense encourage les insectes à visiter les fleurs, cependant, qu’il y ait récompense ou non, cela ne change pas grand-chose pour la fleur du moment que l’insecte propage le pollen.
Comme la synthèse et la production du nectar sont énergétiquement coûteuses, la plante a tout intérêt à réduire ses coûts en trichant. C’est probablement la raison pour laquelle de nombreuses plantes produisent des fleurs chargées en nectar et d’autres sans récompense (« vides ») (Gilbert et al., 1991).
En d’autres termes, ces fleurs proposent un message « malhonnête » par rapport à leur statut de récompense. Les fleurs « malhonnêtes » offrent des avantages en termes de réduction des coûts énergétiques : elles permettent la visite et la fécondation de la plante sans recours à la production de récompenses, du moins auprès des pollinisateurs ignorants ou ne sachant pas distinguer des fleurs avec et sans nectar (figure 7).
Cependant, cette stratégie peut être perdante à partir du moment où la proportion d’insectes pollinisateurs ayant la capacité de détecter la supercherie devient trop importante. Dans ce cas, la fréquence des visites et la fécondation diminueront significativement.
La malhonnêteté ne paie donc que si elle est pratiquée rarement…
Les fleurs indiquent aux pollinisateurs, de manière « honnête », lorsque les réserves en nectar sont épuisées suite à la fécondation. Ce message passe par un changement de couleur et d’odeur (Ito et al. 2021)
Un modèle mathématique a montré que le système de message « honnête » et « malhonnête » n’était stable sur le plan évolutif que si les plantes produisent dans certaines proportions des fleurs avec et sans récompense, et si les visites sont réalisées par une une communauté hétérogène de pollinisateurs informés ou ignorants (Ito et al. 2021).
Dans leur modèle, l’équipe d’Ito (2021) montre que plante et pollinisateurs sachant détecter les signaux électrostatiques sont gagnants, tandis que les pollinisateurs ne pouvant pas le faire n’étaient pas été affectés car n’ayant pas perçu le signal.
Évolution des systèmes électrostatiques de marquage floral
Pour un insecte pollinisateur, l’utilisation d’un système de signalisation reposant sur des charges d’électricité statique n’est certainement pas ou peu coûteux. L’information consiste uniquement à un transfert de charges électriques avec la fleur. Comme expliqué en introduction, la quantité d’énergie électrique impliquée est infime et régénérée durant le vol de l’insecte vers une nouvelle fleur.
Le principal coût est donc supporté par la plante. Bien que l’acquisition de la charge électrique statique ne soit probablement pas coûteuse en elle-même, l’électricité déposée sera dissipée rapidement et la remise en forme du signal électrique est quant à lui coûteux. La période transitoire de recharge réduit les chances de la fleur d’être fécondée par des pollinisateurs comprenant que la fleur vient d’être visitée.
Pour contrer cet inconvénient, l’intérêt de la plante est de diminuer au maximum la durée de recharge du signal électrique. Il a été démontré que les plantes avaient la possibilité d’influencer la durée de recharge du signal en ajustant la conductance (capacité d’un corps à laisser passer une certaine quantité de courant lorsqu’il est soumis à une différence de potentiel) entre le système vasculaire et les tissus de la fleur.
De manière surprenante, il peut être bénéfique pour la plante de porter une marque électrostatique pour imposer au pollinisateur de se spécialiser envers une seule espèce (en effet, pour maximiser la fécondation, il doit visiter des fleurs de la même espèce, au moins durant une certaine période).
Stuart Reynolds suggère ainsi que ce mécanisme de marquage électrostatique des fleurs augmenterait l’efficacité de la pollinisation (intérêt de la plante) et la rentabilité de la collecte de la nourriture (intérêt de l’insecte) dans un environnement majoritairement composé d’une seule espèce végétale.
Pour tester cette hypothèse, l’équipe de Clarke (2021) a mené des expérimentations sur des fleurs de Pétunia. Il s’avère que la marque électrique induite par la visite d’un Bourdon terrestre (B. terrestris) dure environ 100 s (soit un peu moins de 2 min). Cette période semble correspondre au temps moyen entre deux passages successifs de Bourdons dans une même fleur. Le signal empêcherait donc l’insecte de visiter deux fois la même fleur à la suite (figure 8).
Des observations identiques sur des abeilles du genre Anthophora sp. butinant des fleurs de la Vipérine commune (Anchuse strigose) ont permis d’estimer des intervalles de temps moyen entre deux visites de fleurs à 3,25 minutes (Kadmon, 1992).
La durée de recharge du signal électrostatique peut également être corrélée avec le temps nécessaire à la fleur pour se remplir à nouveau en nectar. Ce paramètre varie selon les espèces (luo et al., 2017). Certaines fleurs de petite taille pourront se remplir plus rapidement que d’autres fleurs de plus grande taille ou plus complexes comme Impatiens biflora.
Enfin, la quantité en nectar proposée pourrait également être corrélée avec le temps nécessaire pour qu’un pollinisateur féconde la fleur. Pour maintenir l’insecte plus longtemps dans la corolle, la plante a intérêt à fournir davantage de nectar qui demandera à l’insecte plus de temps pour le collecter
À l’heure actuelle, il n’existe pas données sur la durée de vie des signaux électrostatiques des fleurs autre que celles sur de Pétunia.
Affaire à suivre !
Perspectives
Il est fréquent qu’à la suite d’avancées ou d’observations inédites dans un domaine scientifique, comme le fait que les insectes puissent détecter et interpréter des signaux d’électricité statique, de nouveaux axes de recherches soient explorés par les scientifiques.
Autre axe de recherche, le rôle des charges électrostatiques dans la capture d’insectes par les araignées tisseuses de toile (Araneidae) (Ortega-Jimenez et al. 2013 : Vollrath et al., 2013).
Approfondir les connaissances sur les interactions entre les fleurs et les pollinisateurs peut être intéressant dans la lutte contre les insectes ravageurs (Toyoda, 2020) mais aussi pour améliorer la pollinisation des cultures (Khatawkar et al, 2021).
Des recherches portent également sur l’impact des câbles électrique à haute tension ou sur les radiofréquences des téléphones portables sur l’activité des pollinisateurs (Vanbergen et al., 2019 ; Balmori, 2021). Pour l’instant, il n’existe aucune preuve concluante de ces impacts sur les insectes (Schmiedchen et al., 2018) .
Bibliographie
- Armbruster W.C. (2001) : Evolution of floral form : electrostatic forces, pollination, and adaptative compromise. New Phytologist, 152:181-186 (lien)
- Bowker G. & Crenshaw H.C. (2007) : Electrostatic forces in wind-pollination-Part 1 : Measurement of the electrostatic charge on pollen. Atmospheric Environment, 41(8):1587-1595 (lien)
- Clarke D. ; Whitney H. ; Sutton G. & Robert D. (2013) : Detection and learning of floral electric fields by bumblebees. Science, 340:66-69 (lien)
- Clarke D. ; Morley E. & Robert D. (2017) : The bee, the flower and the electric field : electric ecology and aerial electroreception. Journal of Comp Physiol A, 203:737-748 (lien)
- Corbet S.A. ; James B. & Eisikowitch D. (1982) : Are electrostatic forces involved in pollen transfer ? Plant, Cell & Environment, 5:125-129 (lien)
- England S.J. & Robert D. (2021) : The ecology of electricity and electroreception. Biological reviews, 97:383-413 (lien)
- Gilbert F.S. ; Haines N. & Dickson K. (1991) : Empty flowers. Functional Ecology, 5:29-39 (lien)
- Giurfa M. & Nunez A. (1992) : Honeybees mark with scent reject recently visited flowers. Oecologia, 89:113-117 (lien)
- Heinrich B. (1976) : Heat exchange in relation to blood flow between thorax and abdomen in bumblebees. Journal of Experimental Biology, 64:561-585 (lien)
- Ito K. Suzuki M.F. & Mochizuki K. (2021) : Evolution of honest reward signal in flowers. Proceedings of the Royal Society, B288: 20202848 (lien)
- Khan S. A. et al. (2021) : Electric field detection as floral cue in hoverfly pollination. Scientific Reports, 11:18782 (lien)
- Khatawkar D.S. ; James S.P. & Dhalin D. (2021) : Role of electrostatics in artificial pollination and future agriculture. Current Science, 120:484-491 (lien)
- Leonard A.S. & Papaj D.R. (2011) : « X » marks the spot : the possible benefits of nectar guides to bees and plants. Functional Ecology, 25:1293-1301 (lien)
- Luo E.Y. (2014) : Stimulation of flower nectar replenishment by removal: A survey of eleven animal-pollinated plant species. Journal of Pollination Ecology, 12(7):52-62 (lien)
- Palmer R.A. ; Chenchiah I. & Robert D. (2021) : Analysis of aerodynamic and electrostatic sensing in mechanoreceptor arthropod hairs. Journal of Theoretical Biology, 530:110871 (lien)
- Sutton G. ; Clarke D. ; Morley E.L. & Robert D. (2016) : Mechanosensory hairs in bumblebees (Bombus terrestris) detect weak electric fields. Proceedings of the National Academy of Sciences, 113:7261-7265 (lien)
- Toyoda H. (2020) : Insect Physical control: Electric field-based Pest Management Approach. Insects, 11(8):480 (lien)
- Vaknin Y. et al. (2000) : The role of electrostatic forces in pollination. Plant Systematics and Evolution, 222:133-142 (lien)
- Vaknin Y. ; Gan-Mor S. ; Bechar A. ; Ronen B. & Eisikowitch D. (2001) : Are flowers morphologically adapted to take advantage of electrostatic forces in pollination ? New Phytologist, 152:301-306 (lien)
- Yokoi T. & Fujisaki K. (2009) : Recognition of scent marks in solitary bees to avoid previously visited flowers. Ecological Research, 24:803-809 (lien)





