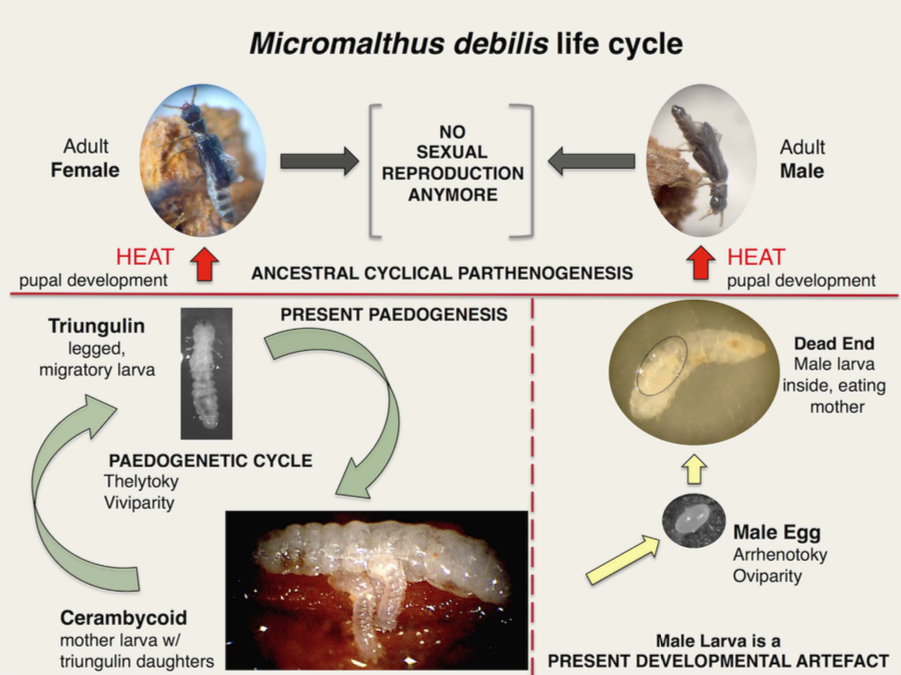
Le cycle de vie incroyable de Micromalthus debilis !
Micromalthus debilis, un Archostemate (plus petit sous-ordre des coléoptères – 50 espèces) d’origine nord-américaine ne semble pas avoir évolué depuis plusieurs dizaines de millions d’années : il est l’unique représentant vivant connu de la famille des Micromalthidés.

Bien que l’espèce affiche des caractères morphologiques archaïques, son cycle de développement est quant à lui à la fois complexe et singulier comme a pu le révéler l’entomologiste américain Herbert S. Barber (1882-1950) – que ses observations en la matière mirent en doute sa crédibilité -. Depuis, seulement cinq expérimentations ont été réalisées sur cette espèce en cent ans.
Le cycle de M. debilis combine à la fois de l’haplodiploïdie, de l’uni-oviparité (un oeuf mâle pondu à la fois), de la poly-viviparité (plusieurs larves femelles sont délivrées à chaque fois) et de la matriphagie interne et externe (consommation de la mère par la descendance).
Tout d’abord, les populations de M. debilis sont exclusivement constituées de larves femelles (néoténie*) qui, chose surprenante, ne donnent naissance à leur tour qu’à des larves également femelles sans passer par le stade adulte. Ce mode de reproduction est appelé parthénogenèse thélytoque (lien) ou en encore pédogenèse. Il existe un dimorphisme entre les stades larvaires : au premier stade elles sont actives et très mobiles (triangulins – 1mm) ; et apodes au second (cérambycoïdes – 3,3mm).
* Néoténie : absence de la phase adulte, le cas le plus connu concerne une sorte de salamandre mexicaine, l’axolotl (Ambystoma mexicana). Les individus adultes possèdent des organes larvaires comme des branchies externes par exemple.


Encore plus surprenant, lorsque les conditions sont défavorables (si le bois dans lequel les larves se développent sèche trop par exemple), la larve au second stade a deux options : 1) Se nymphoses et donner un adulte ailé femelle, 2) Muer une nouvelle fois et livrer une larve de troisième stade également pédogénétique. Cette larve acquière alors la capacité de pondre par parthénogenèse un oeuf unique (arrhénotoque), mâle, puis sombre dans une véritable léthargie. De cet oeuf éclos une larve d’un autre type (curculionide), mâle bien sûr, très différente de la larve femelle : aussitôt, elle enfonce sa tête dans l’orifice génital de sa mère endormie pour… s’en nourrir (matriphagie). Le festin dure une semaine avant que la larve ne se nymphose et ne livre un mâle ailé. Il est possible que cette larve conduite à la matriphagie car elle n’est pas dotée des bactéries symbiotes permettant de digérer le bois.
Les adultes sont très rarement observés, que ce soit dans l’environnement ou en laboratoire. Des études récentes, menées par l’Anglais M. Alejandra Perotti et son équipe, ont mis en évidence que les adultes pouvaient apparaître, in vitro, lorsque des larves cérambycoïdes étaient soumises à des chocs thermiques. Ces chercheurs suggèrent que cet insecte ait quasiment perdu la capacité à se métamorphoser suite à des mutations au niveau de gènes régulateurs clés régulant cette transformation. Le passage à l’état adulte est particulièrement rare car les scientifiques n’ont pu obtenir qu’une seule femelle pour 650 larves et un mâle pour 10 000 larves cérambycoïdes !
Le fait que les mâles soient devenus anecdotiques dans les populations pout s’expliquer de trois manières : 1) Une diminution de la probabilité d’accouplements par un rejets des femelles pour les mâles apparentés et une faible capacité de dispersion de ces derniers, 2) Une compétition pour les ressources locales, 3) La progéniture d’un sexe augmente la fécondité et la valeur reproductrice (fitness) d’un des parents, ici, les mâles tuent et se développent au détriment des femelles, renforçant ainsi le biais du sex-ratio en faveur des femelles. Cependant, les dernières études ne permettent pas de démontrer et de confirmer l’une de ces théories.

D’après l’équipe de Perotti, le biais en faveur des femelles pourrait également s’expliquer par la présence d’un cycle haplodiploïdique. En effet, les populations de nombreuses espèces haplodiploïdes, sociales ou non, comme les guêpes parasitoïdes (lien), possèdent un sex-ratio biaisais en faveur des femelles. Des espèces parthénogénétiques haplodiploïdiques comme le sont Micromalthus debilis, les guêpes Cinipidae et les mouches Cecidomyidae, présentent des cycles similaires avec un biais en faveur des femelles, bien qu’elles soient non apparentées. Cependant, peu d’études se sont intéressées aujourd’hui à ces cycles biologiques.

L’étude de Perotti a confirmé expérimentalement que les adultes sont physiologiquement dans l’incapacité de se reproduire, cette perte étant probablement survenue au cours du temps par dégénération des rares adultes. Bien que des cas de mâles stériles chez d’autres espèces aient pu être décrits, le fait que chez Micromalthus debilis des femelles le soient également reste sans précédent d’après P.D.N. Hebert (1987). Le « coût » biologique que représente la « fabrication » de ces individus biologiquement inutiles expliquerait que l’essentiel du cycle ait évolué vers la parthénogenèse. Les adultes « fantômes » seraient ainsi des reliquats d’un cycle biologique ancestral sexué devenu aujourd’hui simplement vestigial.
Le processus à l’origine de la féminisation des populations peut également faire intervenir des bactéries endosymbiotiques (Rickettsia bellii), détectées chez toutes les larves analysées, connues pour modifier le sex-ratio de la population et induire une « féminisation » du cycle. Si ces bactéries rétablissent la diploïde et donc la formation de femelles, la chaleur permettrait de neutraliser ou de tuer les Rickettsia induisant la formation de mâles haploïdes. Aucun symbiote n’a en effet été détecté chez les adultes.
Quoiqu’il en soit, fort de cette biologie singulière, ce coléoptère « fossile-vivant » minuscule colonise depuis des temps immémoriaux les milieux les plus étonnants, comme les étais des galeries de mines ou les traverses de voie ferrée, en association avec des moisissures. Il a été ainsi introduit dans quasiment toutes les régions du globe.
Source :
- Perotti M.A. ; Young D.K. & Braig H.R. (2016) : The gost sex-life of the paedogenetic beetle Micromalthus debilis. Nature-Scientific Reports, 6:27364 (lien)