Introduction et importance des sacs aériens chez les insectes
Par Benoît GILLES
Les sacs aériens sont une partie essentielle du système respiratoire des insectes, une innovation évolutive qui permet à de nombreuses espèces d’exploiter de nouvelles niches écologiques et d’optimiser des fonctions physiologiques comme le vol, la régulation de la flottabilité et la croissance. Ces structures dérivent du système trachéal, qui permet l’apport en oxygène et l’élimination du dioxyde de carbone, des gaz essentiels pour leur métabolisme énergétique. La structure et la fonction des sacs aériens varient grandement d’une espèce à l’autre, et leur développement est influencé par les besoins physiologiques et environnementaux.
Les premières observations des sacs aériens remontent au XVIIIe siècle, mais l’exploration systématique de leur rôle et de leur évolution a commencé avec des études sur les insectes volants tels que Drosophila melanogaster et d’autres arthropodes trachéates (respirant à l’aire libre vs aquatiques comme les crustacés) (Swammerdam, 1737 ; Guha et Kornberg, 2005).
Aujourd’hui, les sacs aériens sont considérés comme des innovations adaptatives dans la respiration des insectes, en particulier pour ceux nécessitant des capacités de vol élevées ou des adaptations spécifiques à leur environnement.
Phylogénie et apparition des sacs aériens
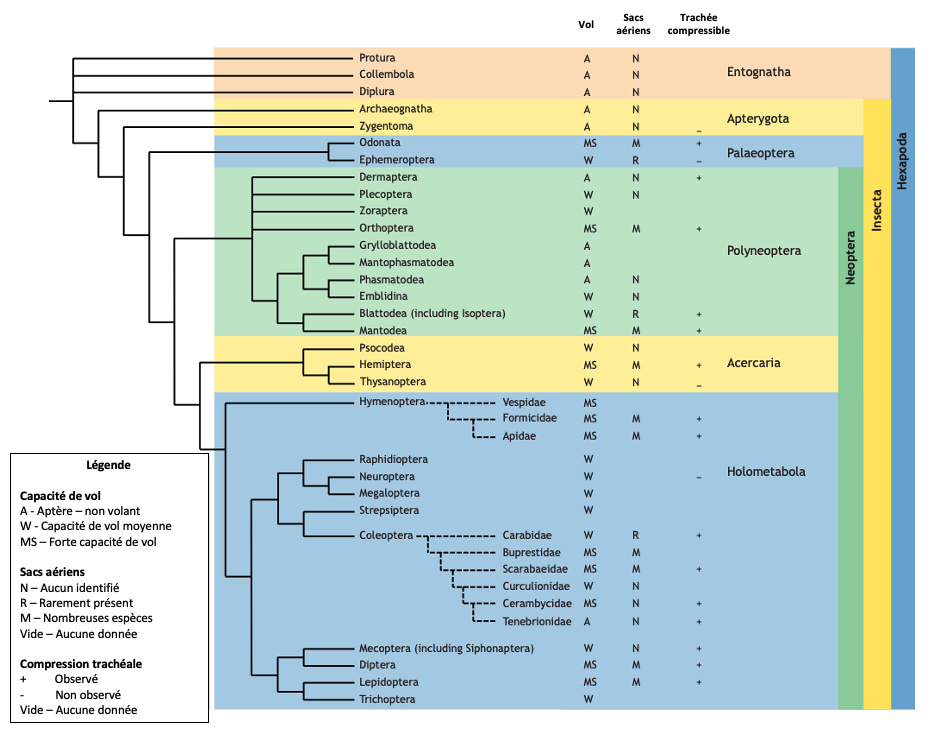
Chez les insectes primitifs, notamment les Paleoptera (Odonates et Éphéméroptères), les sacs aériens sont généralement rares ou limités dans leur développement. Les Odonates, des insectes capables de vol soutenu et puissant, font exception, avec des sacs aériens bien développés chez les adultes. En revanche, les Éphéméroptères, bien que volants, sont des « mauvais » volants et ne possèdent que des structures limitées ressemblant à des sacs aériens dans certains stades larvaires (Herhold et al., 2023).
La présence de sacs aériens est clairement associée aux Neoptera, un groupe d’insectes plus « évolués » qui inclut la majorité des ordres modernes. Parmi eux, les sacs aériens sont particulièrement fréquents dans les groupes ayant développé une capacité de vol élevée, comme les coléoptères, les diptères, les hyménoptères et les lépidoptères (figure 1).
Cette répartition phylogénique des sacs aériens et leur corrélation avec le vol suggèrent que ces organes sont apparus comme une adaptation essentielle pour améliorer l’efficacité respiratoire nécessaire au vol actif (Lee, 1929).
Développement embryologique des sacs aériens
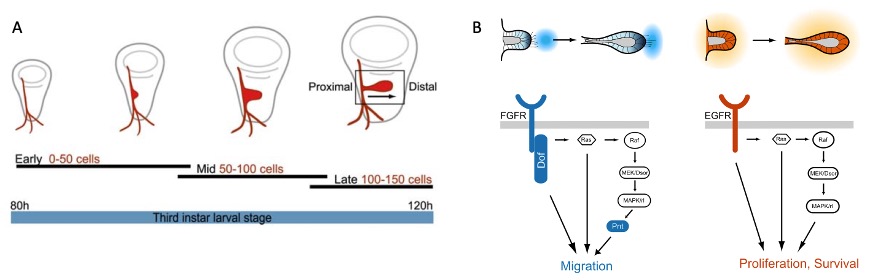
Le développement des sacs aériens est étroitement lié au système trachéal, un réseau de tubes ramifiés qui transporte l’air directement aux tissus pénétrant par des spiracles situés sur l’exosquelette. Le processus de formation des sacs aériens commence au stade larvaire chez des insectes comme Drosophila. Les trachéoblastes, qui sont des cellules progénitrices spécifiques, migrent vers les trachées primaires au cours du développement embryonnaire, puis prolifèrent et se différencient en cellules épithéliales qui formeront les sacs aériens (Cabernard et Affolter, 2005).
Les mécanismes moléculaires sous-jacents à cette formation sont complexes. Par exemple, des récepteurs du facteur de croissance fibroblastique (FGF) régulent la migration des trachéoblastes et la formation des sacs. Ces récepteurs sont activés en réponse à des gradients de FGF exprimés dans les tissus environnants, dirigeant les cellules vers des régions où des structures respiratoires comme les sacs aériens sont nécessaires (Sato et Kornberg, 2002). En parallèle, le facteur de croissance épidermique (EGF) joue un rôle clé dans la régulation de la prolifération cellulaire, contrôlant l’augmentation du nombre de cellules qui formeront les sacs aériens et leur expansion (Cabernard & Affolter, 2005) (figure 2).
Classification morphologique et fonctionnelle des sacs aériens
Les sacs aériens varient considérablement en taille et en structure selon les espèces, et ils peuvent être classés en trois types principaux, selon la présence ou l’absence de taenidies, qui sont des anneaux de renforcement des trachées :
- Sacs taenidiaux : Les taenidies, sont présentes mais plus espacées, permettant une certaine flexibilité. Cette structure est typique chez les insectes ayant besoin d’une certaine rigidité trachéale tout en facilitant la compression et l’expansion (abeilles mellifères – Apis mellifera, criquet migrateur – Locusta migratoria, Odonates par exemple)
- Sacs réticulés : Ils présentent des taenidies irrégulières formant un réseau en croisillons leur conférant une forme irrégulière. Ces sacs ont une capacité de compression accrue, idéale pour les insectes nécessitant des changements rapides de pression interne, comme ceux impliqués dans des cycles de respiration discontinue (Mouche domestique – Musca domestica, Guêpe sociale – Vespa vulgaris, ou le bourdon terrestre – Bombus terrestris par exemple)
- Sacs ponctués : Ces sacs sont les plus compressibles et manquent complètement de taenidies. Ils permettent une grande variabilité de volume et sont souvent trouvés chez les insectes aquatiques ou ceux ayant des régimes respiratoires fluctuants (RRF)
Les RRF se réfèrent aux variations dans les modes de respiration alternant entre des phases actives, de repos ou d’échanges limités. Ces fluctuations s’observent lorsque les besoins en oxygène changent en fonction des activités métaboliques, de l’environnement ou de l’état physiologique de l’insecte.
Faucheux et Sellier (1971) ont proposé cette classification après avoir étudié la structure microscopique des sacs aériens chez plusieurs ordres d’insectes. Ces auteurs ont montré que la morphologie des sacs aériens est fortement influencée par les conditions écologiques dans lesquelles vit l’insecte.
|
Les régimes respiratoires fluctuants (RRF) Un exemple de régime respiratoire fluctuant est le schéma de respiration discontinue (Discontinuous Gas Exchange, ou DGE), fréquemment observé chez les insectes terrestres comme les criquets et les coléoptères. Ce type de respiration comporte trois phases distinctes :
Ce type de régime respiratoire est particulièrement adapté aux environnements où les ressources sont limitées (par exemple, dans des conditions de sécheresse, où la conservation de l’eau est primordiale) ou lorsque l’insecte alterne entre des périodes de repos et d’activité intense, comme le vol ou la prédation. Les sacs aériens, en particulier les sacs réticulés et ponctués, facilitent ces régimes respiratoires fluctuants en permettant des variations de volume qui régulent la pression interne et optimisent les échanges gazeux pendant ces différentes phases. Ces fluctuations de la respiration sont cruciales pour la gestion de l’oxygène et la conservation de l’eau, deux aspects essentiels de la survie dans des environnements difficiles ou lors d’activités métaboliquement exigeantes, comme le vol prolongé ou la recherche de nourriture. |
Faucheux et Sellier (1971) ont proposé cette classification après avoir étudié la structure microscopique des sacs aériens chez plusieurs ordres d’insectes. Ces auteurs ont montré que la morphologie des sacs aériens est fortement influencée par les conditions écologiques dans lesquelles vit l’insecte.
Rôle des sacs aériens dans l’évolution du vol
Les insectes volants ont besoin de maintenir des apports d’oxygène élevés pour alimenter leurs muscles de vol, notamment les muscles thoraciques qui consomment une grande quantité d’énergie lors de la contraction rapide et soutenue nécessaire au vol (figure 3). Les sacs aériens ont probablement évolué en réponse à ces pressions sélectives. Leur fonction principale dans le cadre du vol est de permettre une ventilation par advection (transport d’une quantité d’un élément donné tel que la chaleur, l’énergie interne, un élément chimique par le mouvement (et donc la vitesse) du milieu environnant), augmentant ainsi le flux d’air à travers le système trachéal pour garantir un approvisionnement constant en oxygène (Harrison et al., 2019). Ils permettent également de réduire la densité corporelle de l’insecte, facilitant la manœuvrabilité et diminuant la charge alaire.
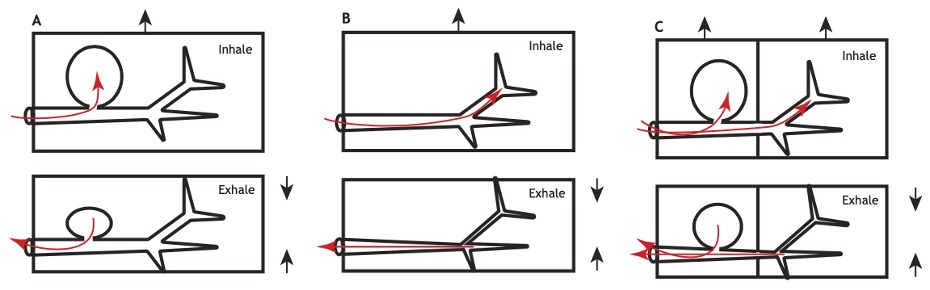
Chez des insectes comme les coléoptères, une forte variabilité est observée dans la présence de sacs aériens au sein même des familles. Par exemple, les coléoptères Scarabaeidae et Buprestidae, qui sont de puissants volants, possèdent des sacs aériens volumineux, tandis que les Carabidae, souvent non volants ou ayant une faible capacité de vol, en sont généralement dépourvus (Dittrich et Wipfler, 2021). Cependant, même parmi les Carabidae, certaines espèces volantes comme les cicendèles ont développé des sacs aériens thoraciques pour soutenir leur capacité de vol, soulignant la forte corrélation entre les sacs aériens et le vol.
Un autre exemple marquant est observé chez les Hyménoptères, en particulier les abeilles et les guêpes. Ces insectes possèdent des sacs aériens thoraciques et abdominaux bien développés qui leur permettent de voler sur de longues distances tout en maintenant une forte activité métabolique. Les abeilles, par exemple, utilisent leurs sacs aériens pour réduire la charge alaire et maximiser l’efficacité du vol lorsqu’elles collectent du nectar et du pollen (Taylor et al., 2010).
Exemples spécifiques
- Chez le scarabée Dicronorrhina derbyana, des images de tomographie micro-CT montrent des sacs aériens massifs entourant les muscles de vol, illustrant leur rôle dans la réduction de la densité corporelle et dans l’amélioration de la maniabilité en vol (Harrison et al., 2019) (figure 4)
- Les orthoptères tels que les criquets ont également des sacs aériens développés au niveau thoracique pour offrir de grandes capacités de vol lors de migrations sur de longues distances (Locusta migratoria). Le volume de ces sacs aériens augmente avec l’âge et la taille corporelle, avec une augmentation significative chez les adultes par rapport aux larves (Greenlee et al., 2009)
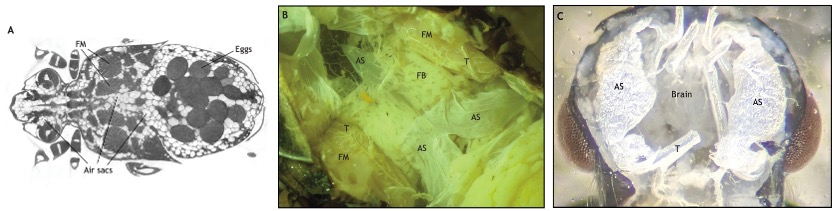
Sacs aériens et stratégies de vol
En plus de leur fonction de soutien à la respiration, les sacs aériens jouent également un rôle important dans la thermorégulation des insectes volants. Chez certains coléoptères, les sacs aériens thoraciques agissent comme des isolants thermiques, aidant à maintenir une température corporelle interne optimale pour le vol endothermique (producteur de chaleur) (Church, 1960). Cela est particulièrement utile pour les insectes volants dans des environnements froids ou lors de vols prolongés, durant lesquels l’évitement de la surchauffe des muscles est crucial.
Les sacs aériens sont également impliqués dans la réduction du coût énergétique du vol. En remplaçant le tissu dense par des cavités remplies d’air, les sacs aériens allègent la structure corporelle de l’insecte, ce qui réduit les besoins énergétiques pour maintenir le vol stationnaire ou prolongé (Newport, 1851). Cependant, il est important de noter que cette réduction de densité peut être contre-productive dans certains cas. Par exemple, une augmentation du volume corporel sans augmentation de masse peut augmenter la traînée aérodynamique lors du vol à grande vitesse (résistance au vent sans gain de puissance musculaire). C’est pourquoi les sacs aériens doivent être soigneusement régulés pour maximiser l’efficacité du vol sans entraîner de pénalités de performance : compression ou expansion (musculaire ou passive) des sacs aériens en fonction des activités et des besoins.
Par exemple, lors d’un vol soutenu chez les bourdons et les abeilles, les sacs aériens thoraciques se dilatent lorsque l’insecte est en vol stationnaire pour maximiser l’oxygénation des muscles de vol, mais ils se compressent partiellement lors de vols rapides pour réduire la traînée (Harrison et al., 2019) (vidéo ci-dessous).
Diversité des sacs aériens et sélection naturelle
La distribution phylogénique des sacs aériens montre clairement qu’ils ont évolué plusieurs fois de manière indépendante chez divers ordres d’insectes.
Cette convergence évolutive illustre l’importance sélective de ces organes pour le vol et d’autres adaptations comme la régulation de la flottabilité.
Les sacs aériens ne sont pas seulement bénéfiques pour les insectes volants, mais ont également permis des innovations évolutives dans la taille corporelle. Par exemple, les coléoptères à « cornes » comme le Rhinocéros (Oryctes nasicornis) ou le Lucane cerf-volant (Lucanus cervus)) ou encore les mouches Diopsidae (vidéo ci-dessous) dont les yeux sont perchés à l’extrémité de longs pédoncules à la manière des requins-marteaux, possèdent des sacs aériens dans leurs cornes, ce qui réduit leur poids sans compromettre leur taille, permettant ainsi la sélection de ces structures sexuelles hypertrophiées (McCullough et Tobalske, 2013).
Extension des pédoncules d’une Diopsidae à l’aide des sacs aériens au niveau de la tête
L’évolution des sacs aériens montre donc un lien fort entre l’optimisation du vol, la réduction de la densité corporelle et les contraintes environnementales. Les insectes ayant besoin de vols prolongés, comme les pollinisateurs ou les insectes migrateurs, ont évolué pour maximiser l’utilisation de ces structures respiratoires complexes, tandis que ceux ayant des besoins respiratoires moindres, comme les insectes terrestres non volants, les ont progressivement perdus.
Compression trachéale : un mécanisme complémentaire
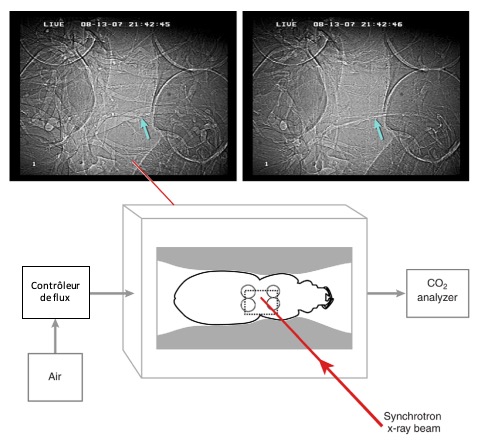
Dans les cas où les sacs aériens sont absents ou sous-développés, les insectes peuvent compenser en utilisant la compression trachéale pour faciliter la ventilation. Ce processus implique une compression des trachées secondaires et tertiaires, souvent à l’aide de mouvements abdominaux. Ce mécanisme permet d’entretenir un flux d’air constant dans le système trachéal sans avoir besoin de structures comme les sacs aériens (Socha et DeCarlo, 2008).
Un exemple notable de ce phénomène s’observe chez les coléoptères carabidés, qui sont généralement des marcheurs terrestres. Ces insectes n’ont pas de sacs aériens développés, mais ils compressent activement leurs trachées pour augmenter le flux d’air pendant l’activité locomotrice intense (Socha et al., 2008) (figure 5). Leurs trachées sont souvent ovales en coupe transversale, permettant une flexibilité accrue lors de la compression, un élément clé pour leur survie dans des environnements nécessitant une mobilité rapide.
Avantages et inconvénients des sacs aériens
Les avantages des sacs aériens sont nombreux, notamment pour les insectes volants. Cependant, ces avantages ont des contreparties importantes. L’un des principaux coûts évolutifs de leur présence est la réduction de la capacité à stocker des réserves d’eau et de nutriments. Ceci est particulièrement crucial dans les environnements secs où les insectes doivent optimiser la rétention d’eau pour survivre (Bradley et al., 1999).
Dans les environnements arides, les insectes ayant de grands sacs aériens peuvent être désavantagés car ils manquent de réserves d’eau suffisantes pour compenser les pertes dues à l’évaporation. En revanche, les insectes plus petits et ceux ayant des structures trachéales plus rigides (moins de sacs aériens) sont mieux équipés pour limiter ces pertes.
Innovations évolutives permises par les sacs aériens
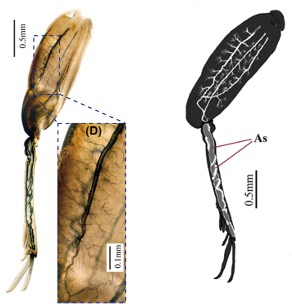
Les tibias d’orthoptères tels que ceux des criquets renferment également des sacs aériens, réduisant la masse des pattes et facilitant ainsi la locomotion (figure 6). Cette adaptation montre que le rôle des sacs aériens n’est pas limité aux fonctions respiratoires, mais qu’ils participent aussi à l’allègement du squelette et à l’optimisation de la locomotion (Ruan et al., 2018).
Adaptations au milieu aquatique et fonction des sacs aériens
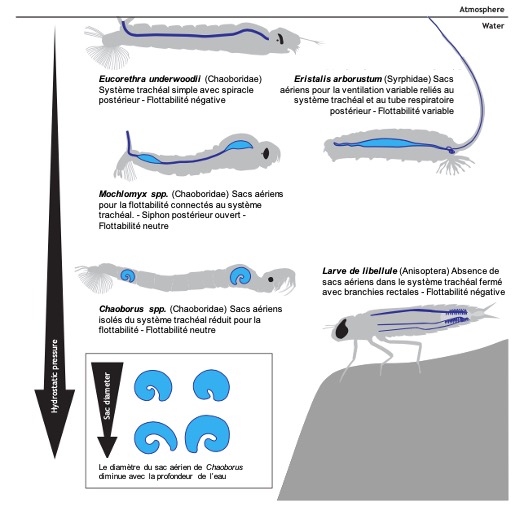
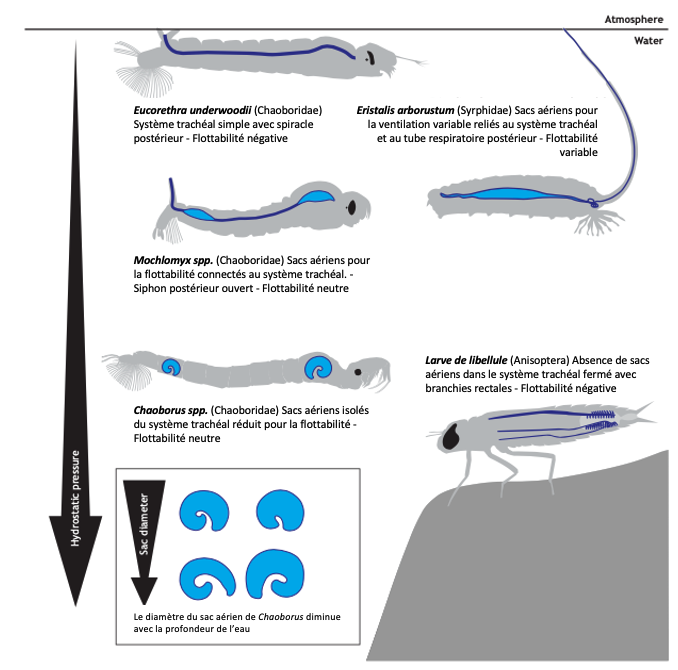
Chez les insectes aquatiques, les sacs aériens sont relativement rares ou limités. En raison de la pression hydrostatique élevée sous l’eau, leur présence peut rendre difficile le maintien de la position sous l’eau. La flottabilité excessive est un problème majeur pour les insectes aquatiques. C’est pourquoi les larves de nombreuses espèces, comme celles des libellules, ont des systèmes trachéaux fermés sans sacs aériens (Tillyard, 1917) (figure 7).
Cependant, certains insectes aquatiques, comme les larves de Chaoborus, ont développé des sacs aériens spécialisés qui leur permettent de réguler leur flottabilité. Ces sacs aériens plus résistants à la pression permettent aux larves de contrôler leur position dans la colonne d’eau, une capacité essentielle pour chasser les zooplanctons et échapper aux prédateurs (McKenzie et al., 2022) (figure 7 et 8).

Régulation de la ventilation trachéale : l’importance des mouvements abdominaux et des cycles de compression
La ventilation trachéale chez les insectes repose sur des mécanismes actifs et passifs. Chez les insectes de grande taille ou à métabolisme élevé, tels que les criquets et les coléoptères, les mouvements rythmiques de l’abdomen ou du thorax provoquent la compression des sacs aériens et des trachées, ce qui génère un flux d’air constant (Westneat et al., 2003).
Les insectes possédant des sacs aériens volumineux, comme certains hyménoptères, montrent des cycles de compression-expansion qui permettent de réguler l’apport d’oxygène et l’élimination du dioxyde de carbone. Ce type de ventilation est particulièrement important lors des périodes de vol intense ou de stress thermique, lorsque la demande en oxygène est maximale.
Conclusion
Les sacs aériens sont des structures adaptatives cruciales qui ont permis aux insectes de développer une grande diversité de stratégies écologiques et physiologiques. De leur rôle dans la respiration et le vol à leurs contributions à la régulation de la flottabilité et à l’évolution de structures complexes (musculatures, appendices hypertrophiés, systèmes de contrôle de la flottabilité, etc.), les sacs aériens ont profondément influencé l’évolution de ces arthropodes.
Leur présence n’est pas seulement une réponse aux besoins respiratoires mais aussi une innovation permettant des stratégies complexes d’adaptation à l’environnement. Cependant, les sacs aériens posent des défis évolutifs, notamment en termes de gestion de l’eau (transpiration, déshydratation, etc.) et de la compression sous pression hydrostatique. Des recherches futures pourraient se concentrer sur l’exploration de la diversité fonctionnelle des sacs aériens en lien avec les stratégies de survie des insectes dans des environnements extrêmes.
Bibliographie
- Bradley, T. J., Williams, A. E. & Rose, M. R. (1999) : Physiological responses to selection for desiccation resistance in Drosophila melanogaster. Am. Zool. 39, 337-345 (lien)
- Cabernard C. & Affolter M. (2005) : Distinct roles for two receptor tyrosine kinases in epithelial branching morphogenesis in Drosophila. Dev. Cell. 9,831-842 (lien)
- Church, N. S. (1960) : Heat loss and the body temperatures of flying insects : II. heat conduction within the body and its loss by radiation and convection. J. Exp. Biol. 37, 186-212 (lien)
- Dittrich, K. & Wipfler, B. (2021) : A review of the hexapod tracheal system with a focus on the apterygote groups. Arthropod. Struct. Dev. 63, 101072 (lien)
- Faucheux, M. J. & Sellier, R. (1971) : L’ultrastructure de l’intima cuticulaire des sacs aeriens chez les Insectes. C. R. Hebd. Séances Acad. Sci. D 272D, 2197-2200
- Greenlee, K. J. et al. (2009) : Synchrotron imaging of the grasshopper tracheal system: morphological components of tracheal hypermetry and the effect of age and stage on abdominal air sac volumes and convection. Am. J. Physiol. Comp. Regul. Integr. Physiol. 297 (lien)
- Guha A. & Kornberg T.B. (2005) : Tracheal branch repopulation precedes induction of the Drosophila dorsal air sac primordium. Dev. Biol. 287, 192-200 (lien)
- Harrison, J. F. et al. (2019) : Physiological responses to gravity in an insect. Proc. Natl. Acad. Sci. USA 117, 2180-2186 (lien)
- Herhold H.W. ; Davis S.R. ; DeGrey SP.P & Grimaldi D.A. (2023) : Comparative anatomy of the insect tracheal system part 1 : introduction, Apterygotes, Paleoptera, Polyneoptera. Bull. Am. Mus. Nat. Hist. 459, 1-184 (lien)
- Lee M.O. (1929) : The function of the air sacs in holopneustic insects. Science 69, 334-335 (lien)
- Mccullough, E. L. & Tobalske, B. W. (2013) : Elaborate horns in a giant rhinoceros beetle incur negligible aerodynamic costs. Proc. R. Soc. B 280, 2013019 (lien)
- Mckenzie, E. K. G., Kwan, G. T., Tresguerres, M. & Matthews, P. G. D. (2022) : A pH-powered mechanochemical engine regulates the buoyancy of Chaoborus midge larvae. Curr. Biol. 32, 927-933.e5 (lien)
- Newport, G. (1851) : On the formation and use of air sacs and dilated trachea in insects. J. Linn. Soc. 20, 419
- Ruan, Y., Li, Y., Zhang, M., Chen, X., Liu, Z., Wang, S. & Jiang, S. (2018) : Visualisation of insect tracheal systems by lactic acid immersion. J. Microsc. 271, 230-236 (lien)
- Sato, M. & Kornberg, T. B. (2002) : FGF is an essential mitogen and chemoattractant for the air sacs of the Drosophila tracheal system. Dev. Cell 3, 195-207 (lien)
- Socha, J. J. & Decarlo, F. (2008) : Use of synchrotron tomography to image naturalistic anatomy in insects. SPIE 2008, 70780A (lien)
- Swammerdam J.J. (1737) : Bybel der Nature. Leyden: Severin
- Taylor, R. A. J., Bauer, L. S., Poland, T. M. & Windell, K. N. (2010) : Flight performance of Agrilus planipennis (Coleoptera: Buprestidae) on a flight mill and in free flight. J. Insect Behav. 23, 128-148 (lien)
- Tillyard, R. J. (1917) : The Biology of Dragonflies (Odonata or Paraneuroptera). Cambridge: Cambridge University press
- Westneat, M. W., Betz, O., Blob, R. W., Fezzaa, K., Cooper, W. J. & Lee, W. K. (2003) : Tracheal respiration in insects visualized with synchrotron X-ray imaging. Science 299, 558-560 (lien)