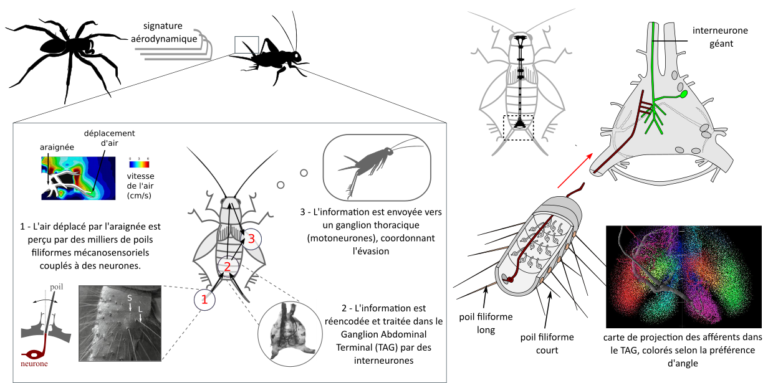
Les pucerons, des insectes passionnants et problématiques (1/2)
Par Maurice Hullé
Petit groupe d’insectes, les pucerons sont cependant parmi les principaux ravageurs des plantes cultivées (lien). En zone tempérée, pratiquement toutes les cultures sont susceptibles d’être affectées. Dans un contexte de réduction des traitements insecticides, il est primordial de bien les connaitre pour mieux les combattre.
Une importante communauté de chercheurs s’intéresse à leur biologie et à leurs impacts agronomiques. Ces insectes fascinent par leur capacité à s’adapter aux changements environnementaux grâce à une série de traits biologiques particuliers qui conditionnent en grande partie leur nuisibilité.
Étudies depuis l’antiquité, les pucerons sont entrés aujourd’hui dans l’ère de la génomique grâce à la description et à l’édition de leurs génomes.
Sexués ou non ?
De la fin de l’hiver à l’automne suivant, les pucerons se multiplient de façon clonale, ne donnant naissance qu’à des femelles parthénogénétiques (lien) vivipares : elles donnent naissance directement à de jeunes larves capables immédiatement de se nourrir. Plus encore ! Une maman puceron porte les embryons de ses filles qui elles-mêmes portent déjà les embryons de ses petites filles. On parle de générations « emboîtées » ce qui constitue un moyen de raccourcir le temps de génération, le développement d’un individu débutant une génération avant sa propre naissance.

Clonalité, viviparité et emboitement des générations permettent un taux de multiplication extrêmement rapide des populations durant la phase parthénogénétique de leur cycle. Ainsi, avec un temps moyen de génération de 2 semaines et une fécondité de quelques dizaines de descendants à chaque génération, une seule femelle donne naissance au cours de sa vie à près de 80 descendantes, qui forment rapidement des colonies très denses de plusieurs dizaines de milliers d’individus. Ce sont ces colonies que l’on observe en manchon sur les tiges des plantes au cours de la belle saison. Tous ces individus sont génétiquement identiques entre eux et à leur aïeule, aux mutations près.
Malgré ce mode de multiplication extrêmement efficace, les pucerons doivent faire face à deux problèmes majeurs. Le premier est d’ordre physiologique : les individus parthénogénétiques vivipares sont sensibles au froid car comme tous les insectes, ils ne régulent pas leur température corporelle. Ils arrêtent de se développer lorsque la température passe en dessous d’un seuil minimal de l’ordre de 4°C selon les espèces et meurent lorsque les températures sont régulièrement négatives. Le deuxième est d’ordre évolutif : comment faire face aux modifications de leur environnement uniquement avec des mutations à fréquence très faible et en absence de la recombinaison génétique liée à la reproduction sexuée ? Les pucerons ont la parade !
Au début de l’automne, percevant la diminution de la photopériode et la baisse des températures, les femelles parthénogénétiques automnales, appelées sexupares, voient leur fonctionnement hormonal modifié et donnent naissance à des descendant clonaux mais sexués, mâles ou femelles fécondables et ovipares. Après l’accouplement permettant l’échange des gamètes, les femelles pondent des œufs qui, eux, résistent au froid. Ces œufs, déposés sur une plante hôte particulière restent au repos tout l’hiver (on dit qu’ils sont diapausants). A la fin de l’hiver, c’est de nouveau une femelle parthénogénétique vivipare qui éclot et qui est à l’origine de nouvelles générations clonales.
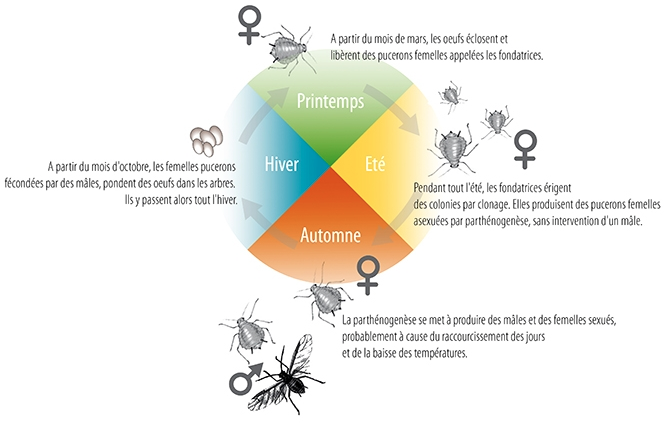
Les pucerons comptent donc parmi les rares animaux à alterner deux modes de reproduction : une phase sexuée durant l’automne et une phase asexuée parthénogénétique durant la belle saison.
Le cycle décrit est le cycle ancestral des pucerons qui, avec quelques variantes, existent depuis 250 millions d’années. On le qualifie d’holocycle et le mode de reproduction associé est la parthénogenèse cyclique. A l’origine, toutes les espèces vivaient sur des plantes arbustives et aujourd’hui de nombreuses espèces de pucerons des arbres pratiquent encore ce type de cycle. C’est le cas par exemple du Puceron lanigère des peupliers Phleomyzus passerinii qui alterne générations sexuée et asexuées sur la même plante-hôte. On parle alors d’holocyclie monœcique (holocyclie sur un seul type de plante-hôte).
Plusieurs grandes modifications de cycle sont apparues au cours de l’histoire évolutive des pucerons. Elles concernent en particulier le passage de la monœcie à l’hétérœcie correspondant à la colonisation des plantes herbacées.

Les pucerons pratiquant l’holocyclie hétérœcique alterne la phase sexuée automnale sur l’arbre (qualifié d’hôte primaire) et la phase estivale d’expansion clonale sur les plantes herbacées secondairement colonisées et qualifiées donc d’hôtes secondaires. C’est le cas de nombreuses espèces ravageuses des plantes cultivées. Citons par exemple Myzus persicae, le Puceron vert du pêcher, dont la reproduction sexuée a lieu principalement sur pêcher et sur d’autres arbres de type Prunus (hôtes primaires) et qui colonise de nombreuses plantes herbacées (photo 1 ci-contre), comme le tournesol, la betterave (vidéo en fin d’article) ou la pomme de terre durant la belle saison (hôtes secondaires), ou bien Rhopalosiphum padi, le Puceron du merisier à grappes qui alterne entre le merisier Prunus padus l’hiver et les céréales du printemps à l’automne.
Une autre étape importante a été l’abandon de l’hôte primaire et l’accomplissement d’un holocycle complet en ne restant que sur hôte secondaire : on parle alors d’holocyclie monœcique sur hôtes secondaires. C’est le cas de Sitobion avenae, le Puceron des épis des céréales, qui accomplit son cycle complet uniquement sur différentes espèces monocotylédones principalement poacées.
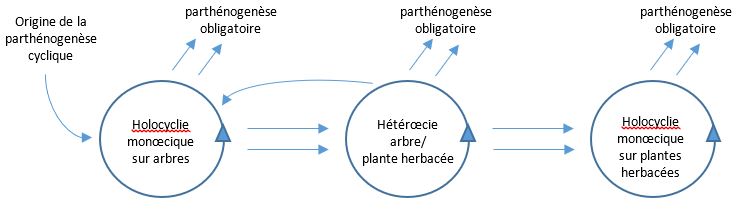
Mais la complexité des cycles biologiques des pucerons ne s’arrête pas là. Une autre transition extrêmement importante est apparue plusieurs fois au cours de leur histoire évolutive. Il s’agit de la perte de la phase sexuée et le maintien toute l’année de la seule reproduction par parthénogenèse. Les aphidologistes parlent d’anholocyclie et le mode de reproduction associé est la parthénogenèse obligatoire.
Seul un petit nombre d’espèces (3%) se maintiennent de façon purement asexuée. Cela leur confère une très forte capacité démographique mais aussi une vulnérabilité importante par rapport au froid hivernal et une faible capacité d’adaptation aux changements environnementaux.

Un grand nombre d’espèces (environ 40% des 4 400 espèces de pucerons) présentent par contre les deux types représentés par des populations holocycliques (qu’on appellera ici sexuées) et des populations anholocycliques, appelées ici asexuées. Ainsi un hiver doux permettra la survie et une bonne multiplication des populations asexuées alors qu’un hiver rigoureux verra ces populations fortement diminuées au profit des populations sexuées qui auront survécu sous forme d’œufs résistants. La coexistence de ces deux types de populations au sein d’une même espèce lui permet de profiter pleinement des hivers doux grâce aux populations asexuées tout en assurant sa survie en cas d’hiver rigoureux.

Cette coexistence est particulièrement avantageuse dans les régions où le climat hivernal fluctue d’une année sur l’autre. Ainsi on observe plutôt des espèces ou des populations asexuées dans les régions à hiver régulièrement doux comme le sud de la France, des populations ou des espèces plutôt ou exclusivement sexuées dans les régions à hiver régulièrement rudes comme l’Est ou le Nord de la France ou de l’Europe et des proportions fluctuantes des deux types de cycle dans les régions intermédiaires à hiver tantôt doux ou tantôt froid comme la Bretagne. De même les cultures sous abri peuvent permettre à des populations asexuées de se maintenir toute l’année.
|
Quels sont les mécanismes permettant la perte de la sexualité ? Quelle est la durée de vie des espèces purement asexuées ? Quels sont les mécanismes régulant la clonalité et la sexualité ? Quelles sont les conséquences écologiques et évolutives de chacun de ces deux modes de reproduction ? Autant de questions de recherche auxquelles une importante communauté scientifique tente d’apporter des réponses. Pour en découvrir davantage (description et identification – pucerons et milieux – pucerons et agricultures – pucerons et recherche), rendez-vous sur Encyclop’Aphid, le site des Aphicionados ! (ou ici) |
Ailés ou aptères ?
Au sein d’un même clone, les pucerons adultes sont soit ailés soit aptères. Les morphes aptères, très féconds, se nourrissent et se multiplient sur une seule plante jusqu’à y produire des colonies denses si les conditions sont favorables. Cela peut entraîner l’épuisement de la plante. Pendant cette phase de multiplication clonale, les morphes ailés (photo 2 ci-dessus), dont la fréquence au sein des colonies augmente avec la densité des populations, assurent alors la dispersion des individus et la colonisation de nouvelles plantes-hôtes (vol de dispersion : D dans le schéma ci-dessous). A leur arrivée sur une nouvelle plante, les pucerons ailés donnent naissance à quelques individus aptères (photo 3 ci-dessus) qui seront, à leur tour, à l’origine de nouvelles colonies. Chez les espèces hétérœciques, les ailés assurent également les passages de l’hôte primaire aux hôtes secondaires au printemps (vol d’émigration : E) puis les vols de retour vers les hôtes primaires à l’automne suivant (vol de rémigration : R).
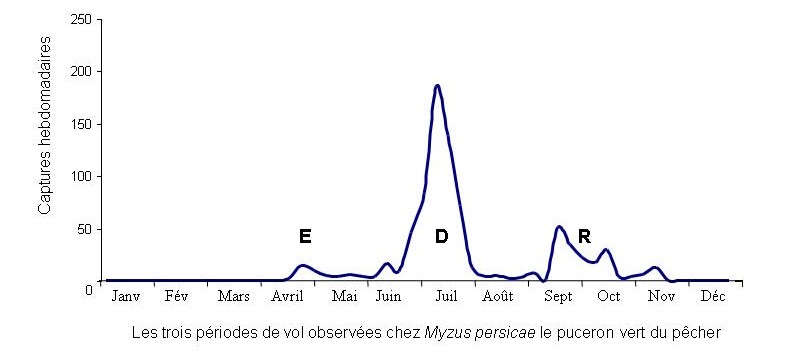
Les différents vols observés au cours de l’année correspondent à des déplacements permettant aux pucerons de rechercher les plantes les mieux adaptées à leur multiplication ou sur lesquelles effectuer la reproduction sexuée.
Toutes ces activités de vols diffèrent selon les espèces et les conditions environnementales. Elles peuvent être détectées par des réseaux de piégeage.
Les formes ailées étant toujours à l’origine de la colonisation des plantes cultivées, leur détection précoce constitue effectivement un outil de prévision des risques phytosanitaires encourus pas les plantes, voire un outil d’aide à la décision d’intervention par les agriculteurs.
En dehors des déplacements par les airs, la dispersion des pucerons est limitée aux déplacements des individus aptères d’une plante vers les plantes situées à proximité immédiate comme ceux d’un même rang dans le cas de la plupart des plantes cultivées, ou aux déplacements non intentionnels comme les transports passifs sur des plantes, des personnes ou des engins agricoles. Les réseaux de piégeage permettent aussi de détecter régulièrement de nouvelles espèces pour les régions où ils sont implantés.
Là aussi les questions de recherche sont nombreuses : elles concernent entre autre la régulation de ce polyphénisme ailé/aptère ou les inductions des formes ailées en réponse à des dangers comme les ennemis naturels.
Les pucerons et leurs plantes hôtes

Les pucerons se nourrissent de la sève élaborée des plantes qui circule dans le phloème, provenant de la photosynthèse. La sève phloémienne est un liquide dont la composition et la qualité nutritionnelle varient au cours des saisons, pauvre en protéines, riche en sucres, limité en acides aminés essentiels non synthétisables par les animaux et quasi exempt de lipides. Autant de barrières évolutives que les pucerons surmontent grâce à des bactéries symbiotiques. La composition protéique et lipidique de la sève oblige le puceron à en ingérer une grande quantité et du coup à rejeter en abondance un surplus en sucre sous forme de miellat (photo 4 ci-contre – vidéo en fin d’article).
Cette ressource sucrée produite par les pucerons est une aubaine pour certains apiculteurs (voir le miel de sapin par exemple) et une nuisance pour certains arboriculteurs ou paysagistes urbains. En effet le miellat attire des micro-organismes dont certains sont pathogènes de plantes (lien) et abîme les produits végétaux comme les fruits en les rendant impropres à la vente.
Les aptitudes de survie et de reproduction des pucerons, dépendent de leurs capacités à localiser et exploiter les plantes dont ils se nourrissent. Environ 5% seulement des espèces de pucerons sont polyphages. Toutes les autres sont plus ou moins spécialisées et colonisent un petit nombre de plantes avec lesquelles ils interagissent. Le puceron doit trouver, reconnaître et s’alimenter sur la plante et la plante se défendre contre les pucerons. Ces interactions plantes-pucerons font l’objet de nombreuses recherches.
Les pucerons ailés, transportés par les courants aériens, peuvent être sensibles à des stimuli visuels comme la couleur jaune ou à des zones de contraste clair/foncé comme les bordures de champs. A plus courtes distances, les pucerons peuvent percevoir les composés volatiles émis par les plantes grâce à des récepteurs olfactifs qu’ils portent sur les antennes, au niveau des rhinaries. Après l’atterrissage sur une plante, le comportement du puceron est affecté par la morphologie et la chimie de la plante.
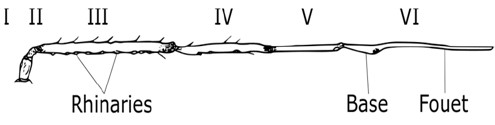
En premier, il peut chercher à rejoindre la face inférieure des feuilles et être gêné dans sa marche par des défenses mécaniques que certaines plantes ont développé telles que les trichomes (photo 5 ci-dessous) ou des productions cireuses. Il peut également rejeter rapidement une plante non-hôte grâce à des chémorécepteurs disposés sur les pattes ou les antennes ou à des piqûres d’essais qui lui permettent de goûter des composés superficiels grâce à des récepteurs gustatifs. De même, il perçoit si la plante est déjà colonisée par des congénères.

Enfin si la plante semble convenir, le puceron commence un processus qui lui permet de confirmer sa reconnaissance, de contrecarrer les défenses de la plante grâce à des effecteurs salivaires puis de s’alimenter longuement le cas échéant. Doté d’un appareil piqueur-suceur, il appuie son rostre à la surface du point de pénétration puis insère ses stylets dans les tissus de la plante le long d’un parcours extracellulaire jusqu’à atteindre, après un certain nombre d’essais, les tubes criblés de la plante où circule la sève phloémienne. La prise de nourriture commence alors. Le puceron s’immobilise et pompe la sève pendant des périodes de plusieurs minutes à plusieurs heures.
La suite de l’article est à retrouver en cliquant sur ce lien
Vidéo
Bibliographie
- Collectif (2007) Pucerons : Les connaître pour mieux les combattre. Dossier Biofutur n°279, 21-57
- Dedryver, C. A., Le Ralec, A., & Fabre, F. (2010) : The conflicting relationships between aphids and men : A review of aphid damage and control strategies. Comptes Rendus Biologies, 333(6-7), 539–553 (lien)
- Dixon, A. F. G. (1998) : Aphid Ecology (2nd ed.). London: Chapman & Hall.
- Hulle M., Coeur d’Acier A., Bankhead-Dronnet S., Harrington R. (2010) : Aphids in the face of global changes, C. R. Biologies (lien)
- Hullé M., Chaubet B., Turpeau E. and Simon J.C. (2019) : Encyclop’Aphid: a website on aphids and their natural enemies. Entomologia generalis (lien)
- Peccoud J. et al. (2010) : Evolutionary history of aphid-plant associations and their role in aphid diversification. C.R. Biologies 333, 474-487 (lien)
- Simon J.C., Stoeckel S., Tagu D. (2010) : Evolutionary and functional insights into reproductive strategies of aphids. C.R. Biologies 333, 488-496 (lien)
Ouvrages sur cette thématique
- Interaction Pucerons-Bactéries-plantes dans la Lutte Chimio Écologique (Jean-Claude Kazadi Mwepu, Odette-Aimée Witanene Sifa & Alain Museu Konga – Editions Académiques – 100 pages – 1 juin 2018)
- Bioécologie des pucerons des agrumes et de leurs ennemis naturels (Karima Benoufella-Kitous & Salaheddine Doumandji – Editions Univ. Européenne – 144 pages – 16 mars 2017)
- Hémiptères de France. de Belgique, du Luxembourg et de Suisse (Romain Garrouste – Editions Delachaux – 320 pages – 27 août 2015)
- Interactions insectes-plantes (Frédéric Marion-Poll & Denis Thiéry – Editions Quae – 750 pages – 5 septembre 2013)