Le système nerveux des insectes
Par Benoît GILLES
Description générale
Identique à celui des autres arthropodes, le système nerveux des insectes, est constitué d’une chaîne nerveuse composée de ganglions reliés entre eux par des cordons nerveux. Ces ganglions regroupent les corps cellulaires (également appelé soma ou péricaryon) de l’ensemble des interneurones et des motoneurones ainsi que les extensions dendritiques et axoniques des neurones (encadré en bas de page). Les axones des motoneurones s’étendent des nerfs périphériques tandis que les axones des neurones sensoriels se dirigent vers les ganglions.
La structuration des ganglions comprend une zone périphérique constituée des corps cellulaires, formant le cortex, et une zone centrale où s’arborisent les neurones qui s’y insèrent (site d’intégration synaptique), appelée la neuropile. Au sein de cette masse de fibres nerveuses, certaines fibres possèdent une orientation commune antéro-postérieure (connectifs), d’autres une orientation transversale reliant la moitié gauche et droite du ganglion (commissures).
Primitivement, chaque segment de l’insecte comportait un ganglion responsable du traitement des informations sensorielles locales provenant des récepteurs musculaires des membres et de la paroi corporelle. Cet arrangement demeure, aujourd’hui, partiellement visible chez de nombreux embryons d’insectes, structure qui se réorganise durant le développement post-embryonnaire. Certains ganglions segmentaires fusionnent avec leurs voisins pour former des neuromères. Le cerveau des insectes correspond à la fusion de plusieurs ganglions de la région céphalique.
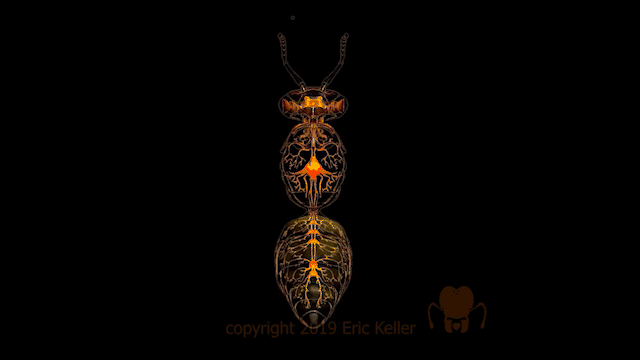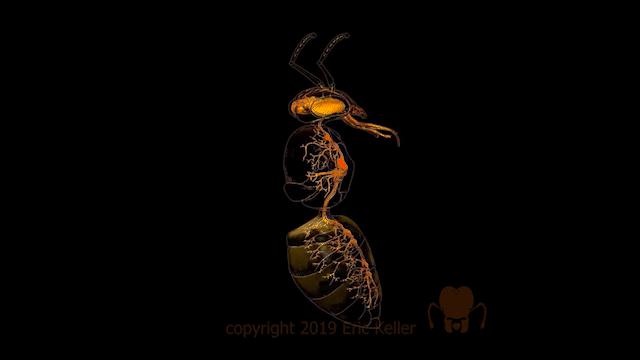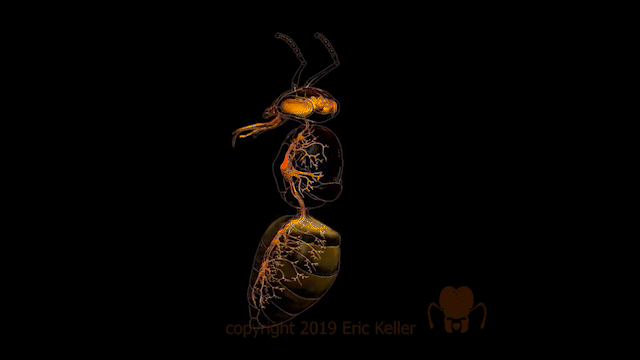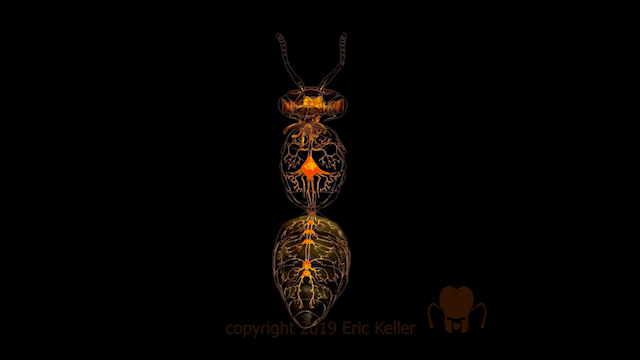
Le système nerveux central est représenté par le cerveau, positionné dorsalement au-dessus de l’intestin antérieur (position dite préorale), et d’une série de ganglions segmentaires situés ventralement.
Dans un axe antéro-postérieur, le premier ganglion de la chaîne ventrale faisant suite au cerveau est, comme son nom l’indique, le ganglion sous-œsophagien. Il résulte de la fusion des neuromères des segments mandibulaire, maxillaire et labial. Chez certains insectes, en particulier les Hyménoptères et les Diptères, le ganglion sous-œsophagien est lui-même fusionné au cerveau, ne laissant qu’un petit conduit à travers lequel passe l’œsophage. Ce ganglion innerve les pièces buccales (lien) (mandibules, maxillaires et labium), les muscles du cou et les glandes salivaires.
Pour tout savoir sur le cerveau des insectes : Le cerveau des insectes
Dans le thorax, il y a généralement trois ganglions. Toutefois, chez certains insectes, deux ou trois d’entre eux sont fusionnés comme chez la mouche Musca (figures 1a et 1b). Le ganglion métathoracique est fusionné avec au moins le premier neuromère abdominal chez presque tous les Ptérygotes (insectes ailés). De chaque côté des ganglions thoraciques diffusent cinq ou six nerfs se ramifiant pour innerver les muscles et les sensilles du thorax et de ses appendices (ailes et pattes) (lien).
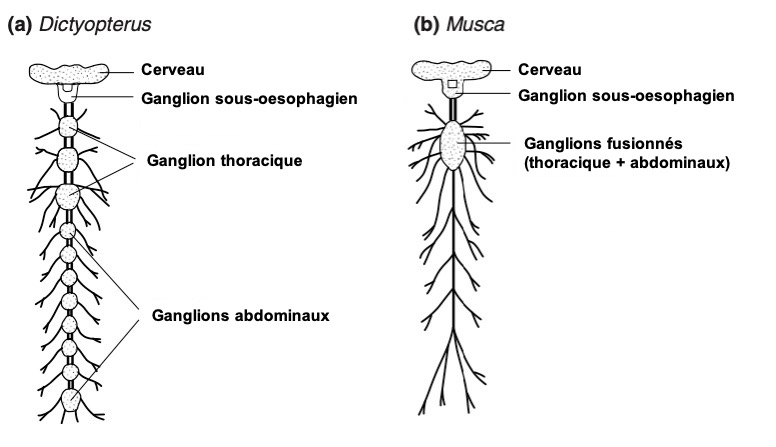
Dans l’abdomen, le plus grand nombre de ganglions abdominaux rencontrés, tant chez des larves et des adultes insectes, est de 8. C’est le cas par exemple chez les Thysanoures ou le mâle Pulex (Siphonaptères). Le dernier ganglion abdominal dérive quant à lui de la fusion des quatre derniers segments abdominaux. Bien que plus nombreux, les ganglions abdominaux sont plus petits et diffusent moins de nerfs que ceux du thorax. De plus, il est constaté une ramification moins importante et diversifiée des nerfs dans l’abdomen. Ces caractéristiques découlent du fait que l’abdomen abrite peu d’organes liés à la motilité nécessitant la présence de neurones moteurs (motoneurones), mais davantage d’organes liés à la digestion et à la reproduction.
Chez certaines espèces, l’ensemble des ganglions abdominaux sont fusionnés comme chez la punaise hématophage Rhodnius et les mouches Cyclorrhapha (en référence à l’ouverture circulaire par laquelle l’adulte sort de la pupe) (figure 2a). En règle générale, un processus de fusion des ganglions se déroule entre la phase larvaire et adulte des espèces holométaboles (lien). Le degré de fusion peut être très variable entre les taxons, même proches phylogénétiquement, entre les mâles et les femelles et selon le statut social chez les abeilles sans dard Melipone.
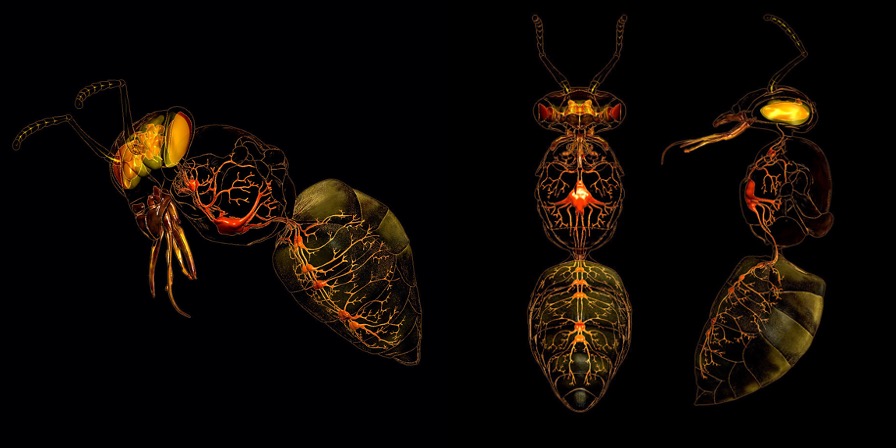
Dans la plupart des cas, les muscles d’un segment sont innervés par des neurones du ganglion du même segment, mais une certaine innervation par des axones provenant des ganglions voisins se produit également (figure 2). Certaines fibres afférentes peuvent également être inter-segmentaires.
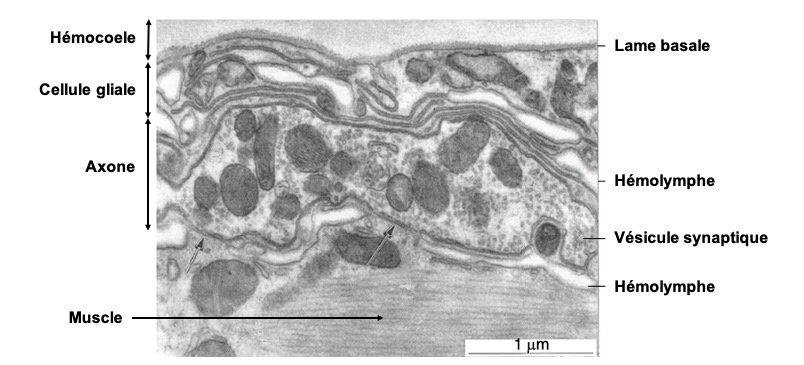
Neurones du système nerveux central
- Neurones moteurs
Les motoneurones interviennent dans le contrôle des muscles. Leur nombre, relativement variable, dépend du nombre d’unités musculaires que contient le segment. Chaque muscle est innervé par 2 à 11 motoneurones. Par exemple, le ganglion métathoracique de la blatte Periplaneta contient 500 neurones.
Chez les insectes, les corps cellulaires des motoneurones sont relativement de grande taille. Par exemple, dans les ganglions métathoraciques de criquets, leur diamètre varie de 20µm à 90µm. Au sein des ganglions, la structuration des motoneurones est symétrique (homologie droite et gauche) et demeure constante entre les individus. Le neurite de chaque motoneurone (prolongement du corps cellulaire d’un neurone : axone ou dendrite) augmente de diamètre en entrant dans la neuropile, au sein de laquelle il donne naissance à une ramification arborescente (figure 3). La forme de cet arbre dendritique est caractéristique de chaque motoneurone, et sa complexité est une indication de celle de ses connexions synaptiques (connexion entre deux neurones) avec les autres neurones. Contrairement aux vertébrés, les insectes possèdent à la fois des motoneurones excitateurs libérant du glutamate et des motoneurones inhibiteurs qui libèrent du GABA (acide γ-aminobutyrique).
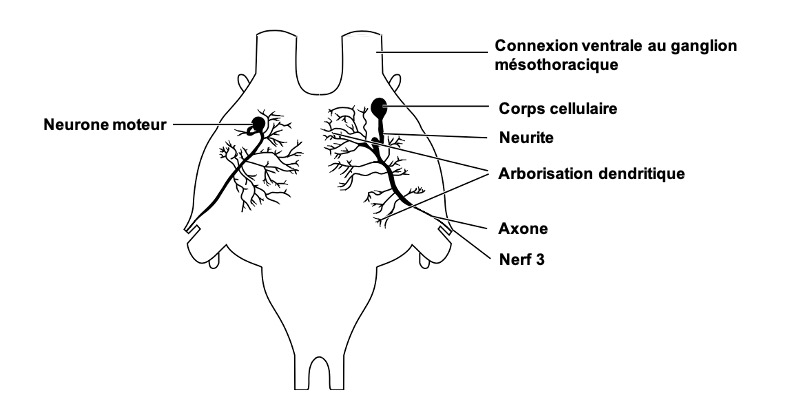
Les motoneurones inhibiteurs diminuent la force de contraction qui peut être produite par un muscle et augmentent son taux de relachement après la contraction. Ils peuvent agir directement sur les fibres musculaires mais peuvent également affecter de manière présynaptique l’activité des motoneurones excitateurs. Les motoneurones inhibiteurs innervent plusieurs muscles, souvent des paires antagonistes, et possèdent des axones particulièrement ramifiés. Par exemple, la patte d’un criquet est innervée par 70 motoneurones excitateurs et par seulement trois motoneurones inhibiteurs.
Les motoneurones excitateurs peuvent être subdivisés en 3 groupes : 1) lente ; 2) intermédiaire et 3) rapide en fonction de la vitesse de contraction musculaire qu’ils entraînent. Chez les insectes, la force et la rapidité de la contraction musculaire dépendent davantage du motoneurone individuel que de l’activité globale d’un grand nombre de neurones fonctionnellement homogènes, comme chez les vertébrés.
- Interneurones
En général, comme leur nom l’indique, les interneurones jouent un rôle d’intermédiaire entre les neurones sensoriels et les neurones moteurs. La plupart des neurones du système nerveux central sont des interneurones. Des connexions synaptiques directes sont toutefois connues pour se produire entre les fibres afférentes (sensorielles) et les motoneurones. Bien qu’apportant une réponse rapide, ces voies monosynaptiques sont rares car cet arrangement entraîne une faible variabilité de la réponse.
Anatomiquement, les interneurones peuvent être divisés en deux groupes : les locaux et les intersegmentaires.
- Les interneurones locaux sont limités à un seul ganglion, pouvant être stimulés ou non. On estime à 1 500 interneurones intraganglionnaires dans le ganglion métathoracique de la blatte Periplaneta, contre 200 interneurones interganglionnaires
- Les interneurones segmentaires transmettent quant à eux des informations le long du cordon nerveux. Les fibres qui transportent des informations des ganglions antérieurs aux ganglions postérieurs sont appelées fibres descendantes, celles qui transmettent des informations au cerveau ou au ganglions antérieurs sont appelées fibres ascendantes. Ces neurones sont responsables de la coordination des activités des différents ganglions et provoquent ainsi la coordination de l’ensemble de l’insecte. Les axones de ces neurones peuvent se ramifier dans un ou plusieurs ganglions voisins ou non, ou transiter à travers des ganglions pour se ramifier uniquement dans des ganglions plus éloignés
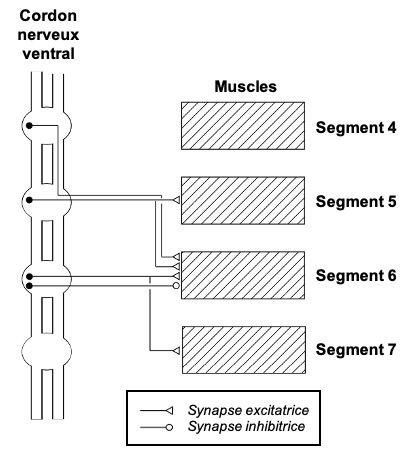
L’un des interneurones intersegmentaires les plus étudiés est le détecteur de mouvement contralatéral (dont l’effet se manifeste du côté opposé au côté atteint) descendant (DCMD) du criquet. Ce neurone, s’étendant du cerveau au ganglion métathoracique, intervient dans le déclenchement de comportements de fuite en présence d’un danger (saut par exemple).
Le déclenchement d’un comportement de fuite fait intervenir un second interneurone entièrement situé dans le cerveau et connecté en partie avec le lobe optique (lobula) : le détecteur de mouvement géant lobula (LGMD). Le LGMD, particulièrement sensible aux stimuli visuels résultant de la détection d’objets mobiles sur la rétine des yeux composés (lien), envoie un signal nerveux au DCMD qui transmet à son tour le signal aux ganglions du thorax : prothoracique, mésothoracique et métathoracique. Ce signal est ensuite orienté vers un certain nombre de neurones excitateurs des muscles du tibia et du neurone inhibiteur commun de la musculature de la patte (figure 4).
Il fournit également une entrée inhibitrice au motoneurone du muscle adducteur coxal. L’activité dans le DCMD, telle que celle qui se produit lors de l’approche d’un objet en collision, est suffisante pour induire des manœuvres de décrochage et de plongée pendant le vol et peut jouer un rôle dans l’initiation d’autres comportement d’évasions et d’évitement des prédateurs.
Au sein d’un grand nombre de taxons comme les blattes, divers orthoptères, les larves de libellules et de drosophiles, voire chez tous les insectes, les cordons nerveux ventraux contiennent des axones appelés fibres géantes.
Chez la blatte par exemple, ces fibres géantes sont au nombre de 6 à 8 avec un diamètre de 20 à 60µm (en comparaison, les axones de la plupart des interneurones font moins de 5µm de diamètre). Le corps cellulaire de ces fibres est situé dans la ganglion abdominal terminal.
Chez la drosophile, les adultes ne possèdent quant eux que deux interneurones géants dont les corps cellulaires se situent dans le cerveau.
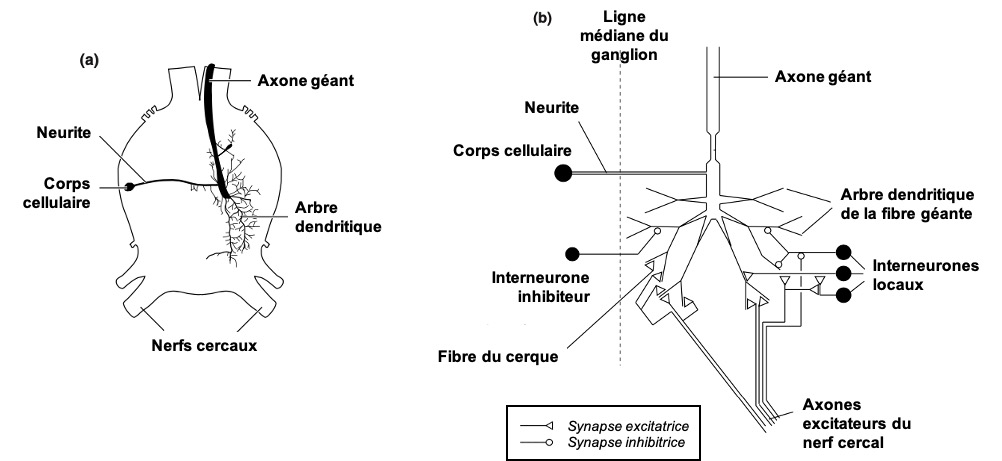
Certaines de ces fibres géantes s’étendent sur des longueurs considérables le long du cordon nerveux. Chez la blatte Periplaneta, les plus grosses fibres s’étendent du ganglion abdominal terminal au ganglion sous-œsophagien, soit sur la longueur du corps de l’insecte. Dans le ganglion abdominal terminal, chaque fibre géante se termine par une arborisation dendritique étendue au sein de laquelle des synapses sont formées avec des fibres nerveuses afférentes provenant du nerf cercal (qui innerve les cerques situés à l’extrémité de l’abdomen) (figures 5 et 6). D’autres fibres géantes se ramifient dans différentes parties du ganglion, suggérant qu’elles reçoivent des informations provenant de différents groupes de sensilles (organes sensoriels) (lien).

Les fibres géantes interviennent dans la transmission rapide d’informations sur de longues distances en raison de leurs taux de conduction élevés, de leur grande taille et de l’absence de synapses. A ce titre, elles jouent également un rôle important dans les mécanismes induisant les comportements de fuite.
- Nerfs périphériques
De chaque ganglion rayonnent des nerfs périphériques constitués d’un grand nombre d’axones indépendants les uns des autres, ne se ramifiant pas et ne formant pas de synapses. Les axones de ces nerfs transmettent l’information provenant des neurones sensoriels périphériques (fibres afférentes) aux ganglions. Ceux distribuant les informations provenant du système nerveux central sont appelés fibres efférentes ou motrices.
Hormis les nerfs antennaires et cercaux constitués uniquement d’axones sensoriels, les nerfs périphériques contiennent à la fois des axones afférents et efférents diffusant symétriquement (homologie) dans un axe droite/gauche. Certains nerfs, comme le petit nerf médian, s’étend d’un ganglion thoracique à l’autre, se ramifiant transversalement aux stigmates et à certains muscles ventilatoires.
NeuroneLes neurones, ou cellules nerveuses, constituent l’unité fonctionnelle du système nerveux. Ce sont des cellules spécialisées pour transmettre des informations à d’autres cellules, nerveuses, musculaires ou encore glandulaires. Chaque cellule nerveuse est constituée d’un corps cellulaire contenant le noyau et le cytoplasme, de dendrites recevant les informations provenant d’autres neurones et d’un axone diffusant les messages nerveux aux autres cellules. Il existe trois types de neurones : 1) les neurones monopolaires, constitués d’un seul axone dont une extrémité joue le rôle de dendrite ; 2) les neurones bipolaires, constitués d’un seul axone et d’une seule dendrite courte généralement non ramifiée recevant les stimuli provenant de l’environnement (organes sensoriels) ; 3) les neurones multipolaires, constitués d’un seul axone et d’une multitudes de dendrites, se rencontrent principalement dans les ganglions du système nerveux sous forme d’interneurones ou de neurones moteurs) (figure 7). Quand un neurone reçoit ou émet un message, il transmet des impulsions électriques le long de l’axone (pouvant mesurer plus d’un mètre). Le message électrique, ou influx nerveux, repose sur l’ouverture et la fermeture de canaux ioniques qui induisent le transfert d’ions à travers la membrane de la cellule, produisant ainsi un courant électrique.  |
Pour tout savoir sur le cerveau des insectes : Le cerveau des insectes
Bibliographie
- The Insects: Structure and Function
(R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)