Les phasmes de Nouvelle-Zélande
Par Morgane Merien
Introduction
Les phasmes appartiennent à l’ordre des Phasmatodea. C’est un ordre très divers, qui comprend plus de 3 000 espèces autour de la planète (Bradler, 2009). Ce sont des insectes phytophages, qui vivent pour la plupart dans des régions au climat tempéré ou tropical. Les phytophages sont des organismes qui se nourrissent d’élément végétaux. Certains phasmes se nourrissent avec un seul type de plantes, alors que d’autres espèces ont un régime plus diversifié.
Le mot « Phasmatodea » vient du grec « Phasma », qui veut dire apparition ou fantôme. Cela se réfère sûrement à leur morphologie particulière ressemblant à une feuille ou un bâton, qui leur donne la possibilité de passer inaperçus dans leur environnement en raison de ce camouflage. Dans d’autres langues, les phasmes sont appelés des « stick insect » en anglais britannique, des « walking stick » en anglais américain ce qui veut dire une « brindille qui marche », ou même « gespenstschrecke » en allemand, ce qui se traduit par « criquet fantôme ».
Les phasmes font partie des groupes d’insectes les plus anciens au monde. Leurs fossiles sont rares, le plus vieil exemple de phyllie remonte à la période Éocène (Wedmann et al., 2007), et le plus vieil exemple de phasme remonte à l’ère Mésozoïque (Tillyard, 1918) (lien).
Le groupe des Phasmatodea a été pendant longtemps considéré comme un sous-ordre du groupe Orthoptera. Ils sont maintenant reconnus comme un ordre à part entière, et un groupe proche des Embioptera. Cependant, les phasmides sont un des rares groupes d’insectes à ne pas avoir une classification basée sur la phylogénétique. Gunther avait classifié les phasmes dans un certain nombre de sous-familles et de tribus, et cette classification avait été largement reconnue. Pourtant, des analyses phylogéniques basées sur des données morphologiques et moléculaires ont plus tard démontré que cette classification n’était pas assez discriminante pour les taxons les plus élevés. Quelques études phylogénétiques ont été réalisées, mais limitées seulement à certains taxons comme le genre Timena en Amérique du Nord, la sous-famille Necrosciinae, et aussi les phasmes de Nouvelle Zélande. Avec ces études, Buckley et al. ont démontré en 2010 que tous les genres de phasmes Néo-Zélandais font partie de la super-famille des phasmes Néo-Calédoniens (lien).
Défense et camouflage
Les phasmes possèdent une large variété de stratégies de défenses pour échapper à leurs prédateurs. Leur stratégie la plus remarquable est leur livrée cryptique. Les phasmes sont des experts en camouflage, ce qui peut devenir frustrant pour toute personne essayant de les chercher dans l’environnement. Le camouflage des phasmes est réalisé par deux méthodes.

La première méthode utilise la couleur du corps, qui se mêle avec la couleur de l’environnent. En anglais, cette forme de camouflage s’appelle « background matching » (Stevens, 2007) (photo 1). La deuxième méthode utilise la forme du corps, qui ressemble à des brindilles qu’on trouverait dans l’environnement. Cette forme de camouflage est appelée mascarade (masquerade en anglais). La mascarade diffère du background matching de façon très importante. Avec le background matching, le système visuel du prédateur n’enregistre pas du tout la présence du phasme.

A l’opposé, avec la mascarade, le système visuel du prédateur repère le phasme mais le classe comme étant une simple brindille au lieu d’un phasme (Troscianko, Skelhorn & Stevens, 2017) (photo 2). Il est maintenant aussi reconnu que le comportement des animaux, et la façon dont cela interagit avec leur apparence, est très important. Il est certain que le comportement des phasmes est important pour renforcer leur camouflage. La façon dont les phasmes oscillent d’un côté à l’autre semble imiter les plantes qui bougent dans le vent (Bian et al., 2016). La position du corps, notamment quand ils tiennent leurs pattes dans le prolongement de leur corps, accentue leur imitation d’un bâton.
Une importante stratégie de défense est l’autotomie. Il s’agit de la capacité d’amputer un membre lorsque ce membre est saisi par un prédateur, leur permettant ainsi de s’échapper rapidement ! C’est aussi très utile, si un membre reste coincé lors d’une mauvaise mue. Si le phasme est une nymphe quand il perd sa patte, il est capable de la régénérer lors des mues suivantes (photo 3 ci-dessus).
Une autre stratégie de défense est la sécrétion de composés chimiques défensifs. Quand elles sont perturbées, certaines espèces de phasmes vaporisent un jet de substances chimiques depuis des glandes tégumentales localisées au niveau du prothorax. Une espèce de phasme Australien est même appelée « peppermint stick insect » (le phasme menthe poivrée) à cause de la substance chimique qu’ils émettent et qui sent comme de la menthe poivrée.
Vidéo d’un peppermint stick insect éjectant la substance chimique :
Un autre mode de défense est la catalepsie. C’est un réflexe d’immobilisation quand le phasme prétend faire le mort, allant même jusqu’à se laisser tomber au sol comme une vraie brindille. Une fois le danger passé, il reprend son activité normale (photo 4 ci-dessous).
Cycle de vie et reproduction
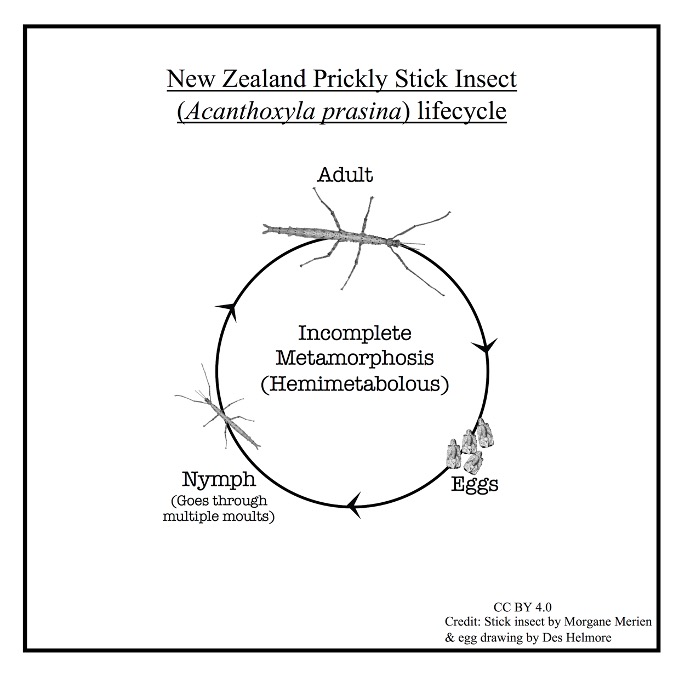
Les phasmes sont des insectes avec un cycle vital de métamorphose incomplète (insectes hémimétaboles). Le cycle de vie se déroule en trois étapes : œuf, nymphe (plusieurs stades) et adulte (figure 1).
En Nouvelle-Zélande, les œufs des phasmes éclosent au printemps et en été (septembre à février), et les nymphes grandissent à travers 7 stades (6 mues) (Stringer, 1970). Les adultes sont communs pendant l’été et l’automne. Comme la majorité des phasmes, ceux de Nouvelle-Zélande sont nocturnes (Bedford, 1978). Pendant la journée, ils restent en position de repos, ou occasionnellement bougent tout doucement pour manger ou accéder à une nouvelle position. A la tombée de la nuit, les phasmes commencent à s’animer, et partent à la recherche de partenaires et de nouvelles plantes à manger. Les mâles détectent les femelles en utilisant des phéromones (signaux chimiques) (Myers et al., 2015).
Les mâles de certaines espèces de phasmes ont une paire de cerques modifiés en forme de pinces, qu’ils utilisent pour rester attachés à la femelle pendant la copulation. Dans certains cas, ils restent même attachés pendant plusieurs heures après avoir fini de copuler, de façon à empêcher d’autres mâles de s’accoupler avec la femelle. La durée de cette surveillance de partenaire semble être influencée par le ratio mâles/femelles observé dans la population (Myers et al., 2015).
La ponte des œufs est similaire pour toutes les espèces Néo-Zélandaises. Les femelles pondent continuellement pendant leur vie adulte, soit par parthénogenèse soit par reproduction sexuelle, et laissent tomber au sol les œufs, où ils resteront dans la litière de feuilles jusqu’à leur éclosion. Les œufs des phasmes sont parfois le seul moyen de différencier les espèces. Toutes les nymphes des espèces Néo-Zélandaises sont vertes quand elles éclosent. C’est à partir de leur première mue que certains phasmes commencent à changer doucement de couleurs, passant généralement du vert au marron.
La majorité des espèces Néo-Zélandaises possède ce polymorphisme de couleurs verte et marron. La durée du développement des œufs peut varier de quelques semaines à quelques mois, en fonction de la période de l’année à laquelle les œufs ont été pondus. Les œufs de certaines espèces plus larges ont aussi tendance à prendre plus de temps pour se développer, par exemple le genre Argosarchus. Dans la mesure où la Nouvelle-Zélande a un climat tempéré, ce long développement associé à une diapause permet aux phasmes de survivre aux conditions les moins favorables, jusqu’à que les températures remontent. La majorité des phasmes adultes meurent avec l’arrivée de l’hiver. Il est parfois rare d’en trouver pendant les mois plus froids, mais ces individus sont normalement assez fins et léthargiques (Salmon, 1991).

Les phasmes de Nouvelle-Zélande
En Nouvelle-Zélande, il y a 23 espèces de phasmes et 10 genres, tous endémiques (Buckley, Attanayake, Nylander, & Bradler, 2010). Ils sont divisés en deux sous-familles : Phasmatinae (tribu : Acanthoxylini) et Pachymorphinae (tribu : Pachymorphinini) (Günther, 1953). Cette classification et identification est basée sur Jewell & Brock (2002) (figure 1).

Toutes les espèces de phasmes Néo-Zélandais ressemblent à des bâtons. Ils n’ont pas d’ailes, et leur couleur varie du vert au marron. Les phasmes sont assez communs en Nouvelle-Zélande, et se trouvent facilement dans les jardins urbains et les forêts natives.
Ils ont de nombreux prédateurs, comprenant des oiseaux endémiques et introduits, des guêpes exotiques, et des espèces de mammifères invasives comme les rats, hermines, hérissons, et opossums.
Dans la langue Māori, le peuple indigène de Nouvelle-Zélande, les phasmes sont appelés Rō ou Whē. Les traditions Māori disent que tous les insectes sont les enfants du dieux Tāne Mahuta, et que par conséquent ils ont droit au respect de tous (Weta de Nouvelle-Zélande). Certaines légendes disent que les phasmes sont des proches parents des mantes religieuses, et que si l’un des deux se pose sur une femme, cela veut dire qu’elle est enceinte et que l’insecte lui-même prédirait le sexe de l’enfant. Dans un autre proverbe, si un phasme tombe sur quelqu’un dans la forêt, cela veut dire qu’il est entré dans un endroit sacré (Trewick, 2007).
Le genre Acanthoxyla contient 8 espèces. Ces espèces sont exceptionnelles car elles ne comportent que des femelles, et se reproduisent par parthénogenèse, une méthode de reproduction asexuée. Acanthoxyla est un genre commun à travers la majorité de la Nouvelle-Zélande, et peut être trouvé sur des plantes communes de jardin comme les roses ou les conifères ornementaux, mais aussi sur des arbres endémiques comme les rimu, les totara et les rata. Ce sont de larges insectes, mesurant à peu près 8 à 11 cm, la plupart de couleur verte ou marron (photo 6). Toutes les espèces ont des épines noires caractéristiques du genre, sauf l’espèce A. inermis qui n’a pas du tout d’épines.
Le genre Argosarchus ne contient qu’une espèce valide. Argosarchus horridus est le plus grand phasme de Nouvelle-Zélande, mesurant entre 15 cm et 20 cm (photo 7). Ce phasme est trouvé dans la majorité de la Nouvelle-Zélande, et fut le premier phasme Néo-Zélandais découvert et décrit en 1846. Comme beaucoup d’autre phasmes Néo-Zélandais, certaines populations ne comprennent que des femelles et se reproduisent par parthénogenèse.

Le genre Clitarchus possède l’espèce la plus commune en Nouvelle-Zélande, Clitarchus hookeri (photo 8). Cette espèce possèdent plusieurs couleurs, variant de vert pomme, à gris lichen et à marron foncé. Ce sont de larges insectes, mesurant près de 11 cm pour les femelles, et 7,4 cm pour les mâles. Les femelles peuvent souvent être confondues avec A. inermis, mais elles sont différenciées grâces à leur cerques, la taille de leur tête et la forme de leurs œufs. Les cerques de C. hookeri sont pointus, et leur tête est moins bulbeuse que A. inermis. Comme dans beaucoup d’autres espèces de phasmes, les mâles sont plus fins et plus petits, ressemblant à des allumettes. C. hookeri a une reproduction avec parthénogenèse facultative, et certaines populations survivent sans mâles. Les deux autres espèces sont limitées à certains habitats de la Nouvelle-Zélande.
Le genre Pseudoclitarchus ne possède qu’une seule espèce, qui n’est trouvée que sur une petite île appelée Three Kings, au Nord de la Nouvelle-Zélande. Les femelles mesurent jusqu’à 7,5 cm, et les mâles 5,6 cm. Il n’y a pas beaucoup d’informations biologiques sur cette espèce (photo 9).

Le genre Tepakiphasma est monotypique. L’espèce Tepakiphasma ngatikuri a été découverte en 2008, puis décrite en 2010 (Buckley & Bradler, 2010). Cette espèce est endémique dans une seule parcelle de forêt la plus au nord de la Nouvelle-Zélande. Tepakiphasma ngatikuri mesure à peu près 10 cm, est de couleur grise et marron, le corps est lisse avec peu de tubérosités. La caractéristique la plus distinctive de cette espèce est l’œuf. L’opercule de l’œuf est perforé et possède une forme conique. C’est une structure qui n’apparaît pas sur les œufs des autres espèces Néo-Zélandaises, mais apparait chez des espèces d’Outre-Mer.
Le genre Asteliaphasma contient deux espèces décrites, et qui ne sont trouvées que dans la moitié Nord de l’île du Nord. Les individus d’Asteliaphasma peuvent mesurer jusqu’à 9 cm, et ont tendance à être assez graciles avec des membres fins. Les œufs sont couverts de fines épines. Les descriptions se rapportant à ces deux espèces ne permettent pas de les identifier de manière fiable (photo 10).

Le genre Micrarchus possède une seule espèce, Micrarchus hystricuelus (photo 11). Cette espèce peut être trouvée dans la partie Sud de l’île du Nord, et la partie Nord de l’île du Sud. Cependant, un certain nombre d’espèces de Micrarchus présentes dans l’île du Sud ne sont pas encore décrites. Les phasmes de M. hystricuelus sont petits, mesurant à peu près 5 à 6 cm pour les femelles et 3,5-4 cm pour les mâles. Ils sont toujours de couleur marron et avec beaucoup d’épines.

Le genre Niveaphasma contient une seule espèce, Niveaphasma annulata. Cette espèce est commune dans la partie Sud de l’île du Sud, mais est complètement absente dans l’île du Nord. Les individus sont de couleur grise ou marron, avec des excroissances distinctes le long du corps et des membres. Ce genre est assez exceptionnel parmi les phasmes, car les individus de cette espèce peuvent être trouvés dans les zones alpines (Dennis et al., 2014).
Le genre Spinotectarchus contient une seule espèce, Spinotectarchus acornutus. Cette espèce est limitée à la partie Nord de l’île du Nord et à quelques îles. Les femelles mesurent à peu près 4,8 cm, et les mâles 3,7 cm (photo 12). Les œufs de cette espèce sont couverts de fines épines, ce qui leur donne une apparence poilue. Cette caractéristique est partagée avec le genre Asteliaphasma, mais leurs organes génitaux sont différents. Des études phylogénétiques ont démontré que ce genre n’est pas apparenté aux autres genres de phasmes Néo-Zélandais (Buckley et al., 2010).
Le genre Tectarchus contient 4 espèces. Elles sont toutes trouvées dans le Nord de l’île du Nord. Les espèces T. huttoni et T. ovobessus sont typiquement de couleur verte, avec quelques rares formes marrons (photo 13). Tectarchus salebrosus et T. semilobatus sont typiquement marron clair ou foncé. La meilleure façon de différencier les espèces de ce genre est de les identifier à partir des œufs. Les phasmes de ce genre sont petits, avec les femelles mesurant entre 5 à 6 cm, et proche de 4,5 cm pour les mâles.
Des phasmes Néo-Zélandais proche de chez nous !
Si vous voulez voir des phasmes Néo-Zélandais depuis la France, vous n’avez pas besoin d’aller très loin! Il suffit juste de traverser la Manche et de vous retrouver dans la région des Cornouailles et du Devon, dans le sud de la Grande-Bretagne. Trois espèces de phasmes Néo-Zélandais y survivent. Il y a Clitarchus hookeri sur l’île de Scilly, et Acanthoxyla geisoveii & Acanthoxyla inermis sur l’île principale. Il y a environ 100 ans, certaines plantes australes comme les fougères étaient très prisées par les britanniques. Un commerce s’est mis en place pour faire venir des plantes de Nouvelle-Zélande (et d’autres endroits du pacifique), avec l’installation d’une pépinière principale située à Truro, dans les Cornouailles, et de deux autres à Paington dans le Devon, et à Tresco dans les îles Scilly. Les œufs, ou même peut être les nymphes, ont dû être transportées avec les plantes depuis l’autre côté de la planète.
Arrivées dans le sud de l’Angleterre, elles ont dû trouver le climat assez agréable et ont pu subsister grâce aussi à la parthénogenèse et à une alimentation variée (Lee, 1993). La présence d’un phasme Néo-Zélandais en Angleterre fut rapportée pour la première fois en 1910. Cela a même fait les nouvelles locales à l’époque ! Depuis, la population de ces deux espèces a continué à se développer et à coloniser de nouveaux habitats. C’est une sorte de gentille invasion que les locaux adorent !
Morgane Merien Je m’appelle Morgane Merien et je suis Doctorante en Sciences biologiques à l’Université d’Auckland en Nouvelle-Zélande. Je suis née en Nouvelle-Calédonie, mais je vis en Nouvelle-Zélande avec ma famille depuis 2006. Cela fait maintenant près de 4 ans que je travaille sur les phasmes de Nouvelle-Zélande. J’ai toujours aimé le monde naturel, surtout en ayant grandi dans des pays ou régions comme la Nouvelle-Calédonie, la Guyane et bien sur la Nouvelle-Zélande. Mon intérêt pour les insectes et les phasmes a pris naissance pendant ma licence de biologie, lorsque j’ai assisté à un cours d’entomologie. C’est à ce moment-là que je suis vraiment tombée amoureuse du monde des insectes. Ce qui m’a attiré, c’est surtout la diversité des espèces, leur comportement et biologie, et leur importance pour notre monde. Pendant ce cours, j’ai su immédiatement que je voulais être entomologiste ! Mon doctorat étudie le camouflage et le polymorphisme des phasmes de Nouvelle-Zélande. La première partie de ma recherche se concentre sur le type de camouflage que les des phasmes utilisent, et comment ils sont perçus par le système visuel de leur prédateur. J’essaie aussi de comprendre comment leur comportement, mouvement et choix de substrat peuvent influencer l’efficacité de leur camouflage. La deuxième partie de mon Doctorat s’intéresse aux couleurs des phasmes, particulièrement le polymorphisme vert-marron retrouvé dans la majorité des espèces néo-zélandaises. Je suis en train de faire des expériences pour déterminer si le choix d’un substrat avec une couleur particulière a une influence sur leur survie. J’espère pouvoir vous communiquer tous ces résultats très prochainement ! |
Bibliographie
- Bedford, G. O. (1978) : Biology and ecology of the Phasmatodea. Annual review of entomology, 23(1), 125-149
- Bian, X., Elgar, M. A., & Peters, R. A. (2016) : The swaying behavior of Extatosoma tiaratum : motion camouflage in a stick insect? Behavioral Ecology, 27(1), 83-92 (lien)
- Bradler, S. (2009) : Phylogeny of the stick and leaf insects (Insecta : Phasmatodea). Species, Phylogeny and Evolution, 2, 3-139
- Buckley, T. R., & Bradler, S. (2010) : Tepakiphasma ngatikuri, a new genus and species of stick insect (Phasmatodea) from the Far North of New Zealand. New Zealand Entomologist, 33(1), 118-126 (lien)
- Buckley, T. R., Attanayake, D., Nylander, J. A., & Bradler, S. (2010) : The phylogenetic placement and biogeographical origins of the New Zealand stick insects (Phasmatodea). Systematic Entomology, 35(2), 207-225 (lien)
- Günther, K. (1953) : « Über die taxonomische Gliederung und die geographische Verbreitung der Insektenordnung der Phasmatodea ». Beiträge zur Entomologie, 3(5), 541–563
- Jewell, T., & Brock, P. D. (2002) : A review of the New Zealand stick insects: new genera and synonymy, keys, and a catalogue. Journal of Orthoptera Research, 189-197 (lien)
- Lee, M. (1993) : A survey into the distribution of the stick insects of Britain. Phasmid Studies, 4(1), 15 (lien)
- Myers, S. S., Buckley, T. R., & Holwell, G. I. (2015) : Mate detection and seasonal variation in stick insect mating behaviour (Phamatodea: Clitarchus hookeri). Behaviour, 152(10), 1325-1348 (lien)
- Salmon, J. T. (1991) : The stick insects of New Zealand. Raupo
- Stevens, M. (2007) : Predator perception and the interrelation between different forms of protective coloration. Proceedings of the Royal Society B: Biological Sciences, 274(1617), 1457-1464 (lien)
- Stringer, I. A. N. (1970) : The nymphal and imaginal stages of the bisexual stick insect Clitarchus hookeri (Phasmidae : Phasminae). New Zealand Entomologist, 4(3), 85-95
- Tillyard, R. J. (1918) : Mesozoic insects of Queensland. No. 3. Odonata and Protodonata. Proceedings of the Linnean Society of New South Wales, 43(3),417-436
- Trewick, S. :’Stick insects’, Te Ara – the Encyclopedia of New Zealand, http://www.TeAra.govt.nz/en/stick-insects/print (accessed 1 October 2020)
- Troscianko J, Skelhorn J, Stevens M. (2017) : Quantifying camouflage : How to predict detectability from appearance. BMC evolutionary biology. 17:7 (lien)
- Wedmann, S., Bradler, S., & Rust, J. (2007) : The first fossil leaf insect : 47 million years of specialized cryptic morphology and behavior. Proceedings of the National Academy of Sciences, 104(2), 565 (lien)