
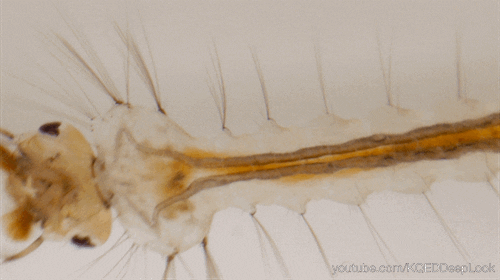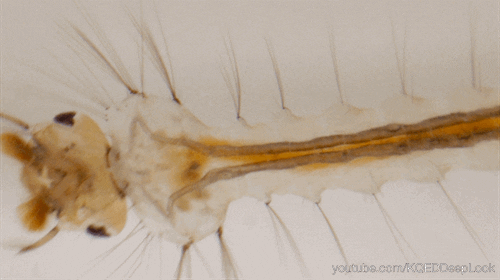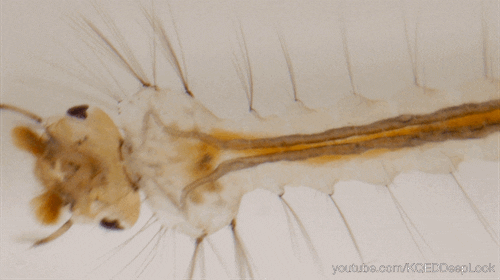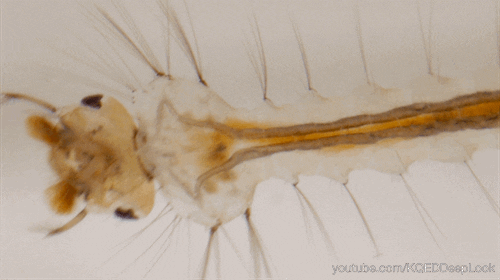
Nombre de vues : 1 285 Par Benoît GILLES Chez les insectes, le système circulatoire se distingue de celui des vertébrés à la fois par sa
Catégorie : Biologie des insectes
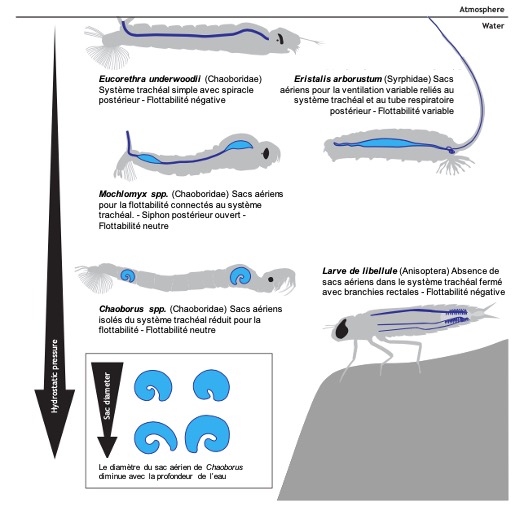
Introduction et importance des sacs aériens chez les insectes
Nombre de vues : 4 540 Par Benoît GILLES Les sacs aériens sont une partie essentielle du système respiratoire des insectes, une innovation évolutive qui permet
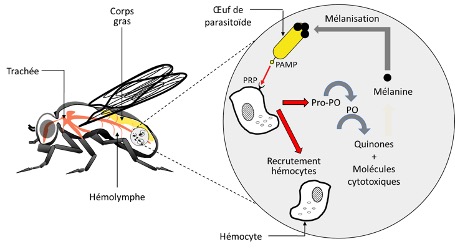
Le système immunitaire des insectes
Nombre de vues : 3 867 Par Fanny Vogelweith Tout au long de leur cycle de vie – allant de l’œuf jusqu’à à la fin de
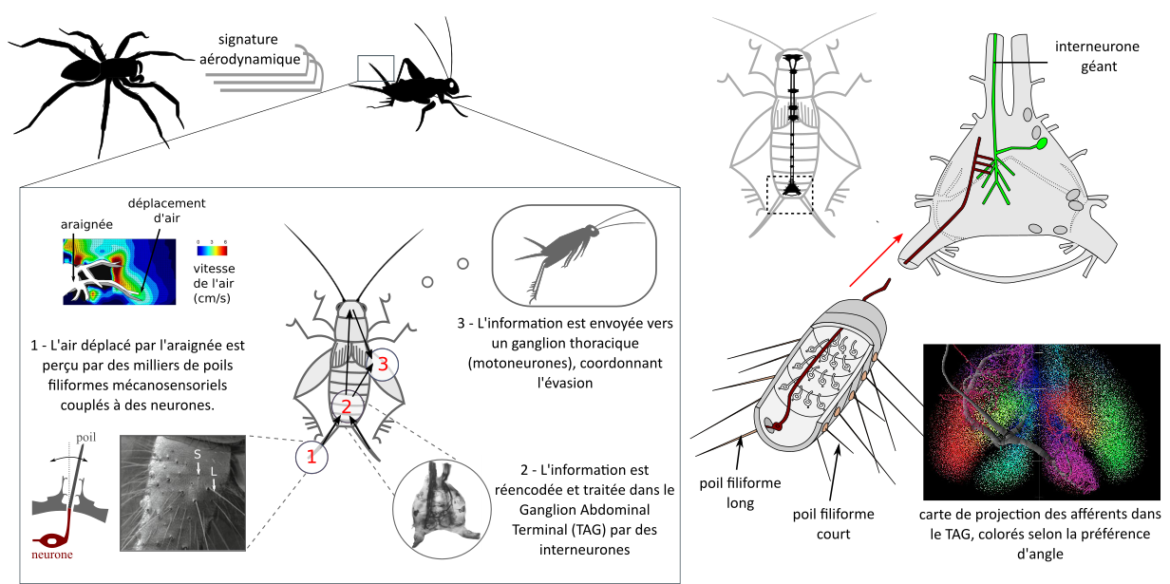
Le guide évasion – Partie 2 : de la perception de stimuli dangereux aux actions motrices
Nombre de vues : 3 901 Par Paul Clémençon Cet article sur la neuroécologie des courses-poursuites est divisé en deux parties. Ne manquez pas la première
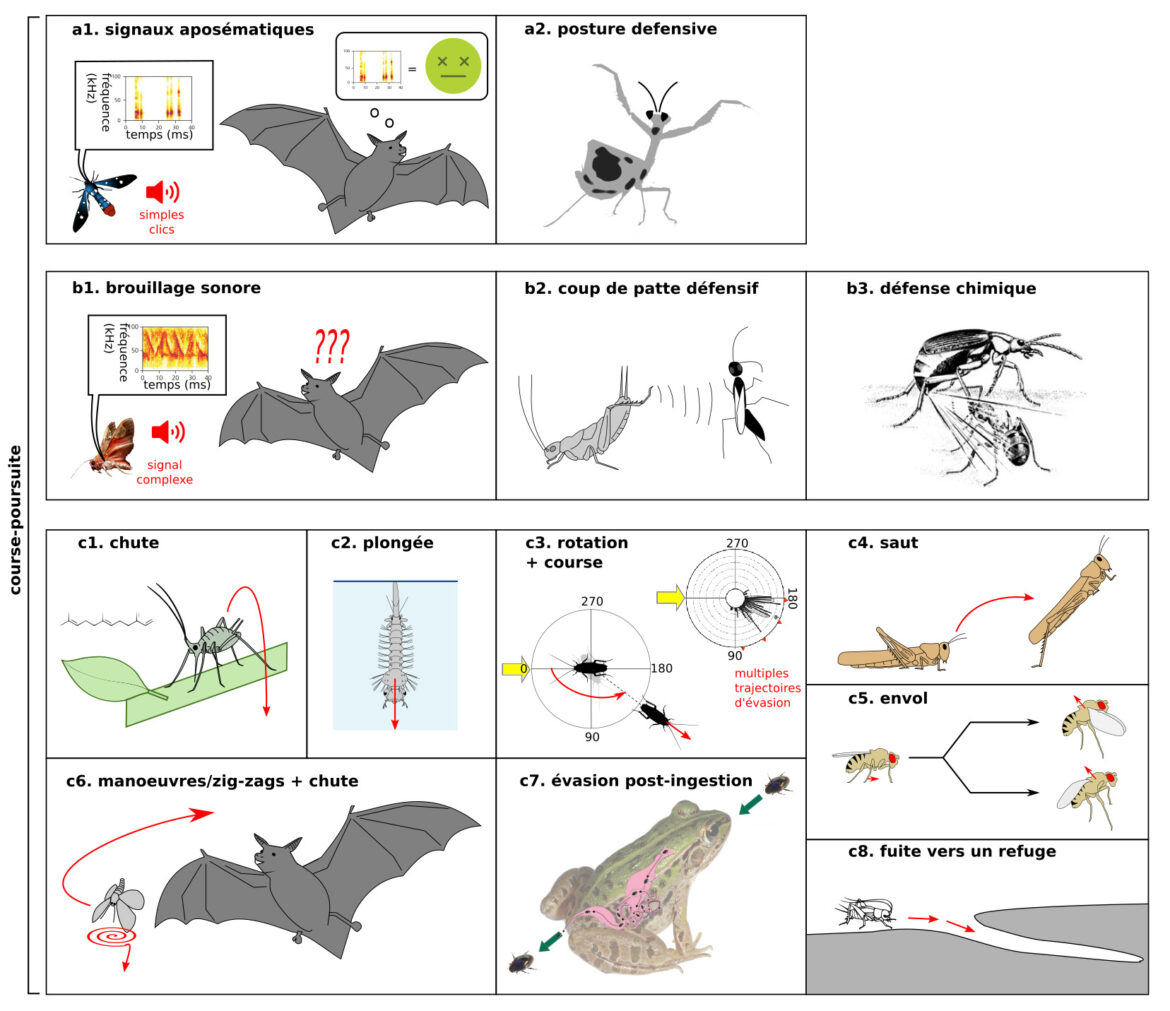
Le guide évasion – Partie 1 : écologie des jeux de cache-cache et de poursuite évasion
Nombre de vues : 3 441 Par Paul Clémençon Cet article est la suite de l’article sur la navigation spatiale chez les insectes : https://planet-vie.ens.fr/thematiques/ecologie/ethologie/s-orienter-vers-la-bonne-route-la-navigation-spatiale-chez-les. Il se
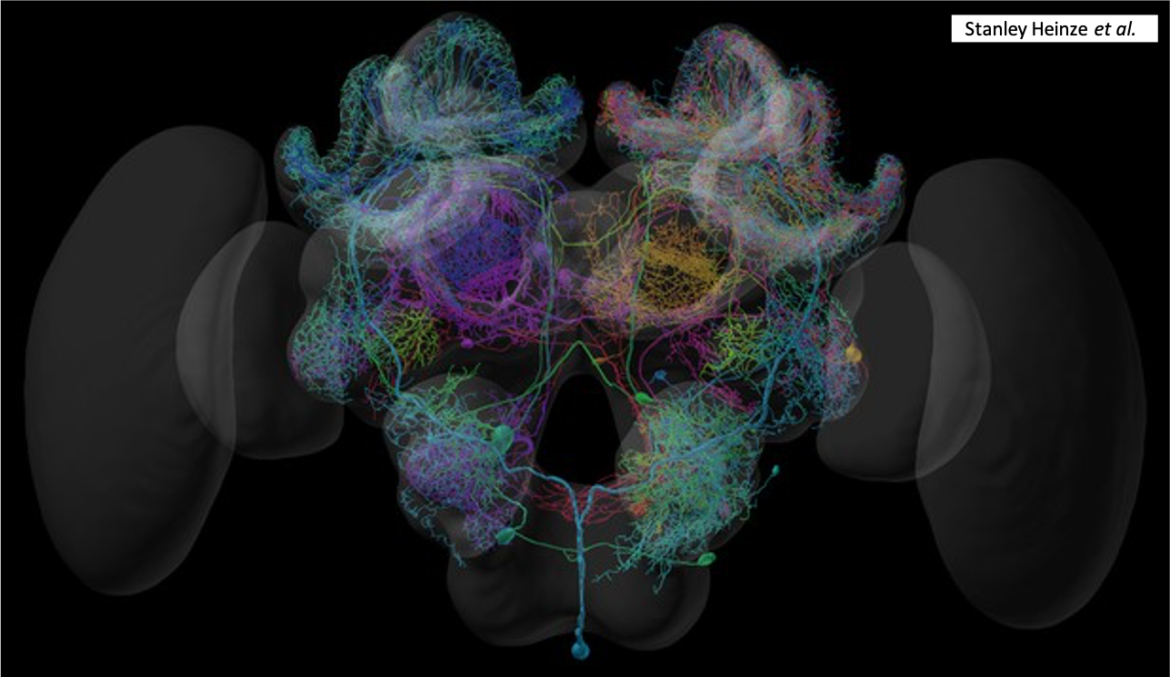
Le cerveau des insectes
Nombre de vues : 7 484 Par Benoît GILLES Que ce soit chez les arthropodes ou les vertébrés, le cerveau constitue le principal centre de contrôle
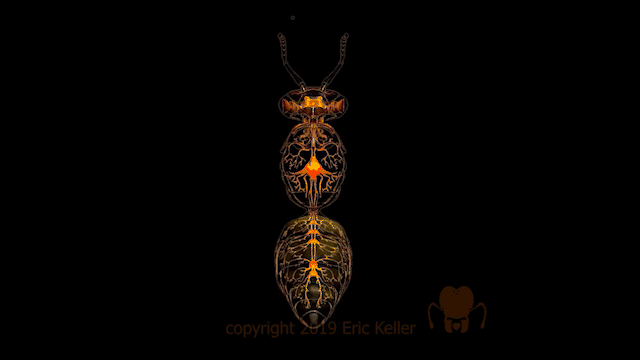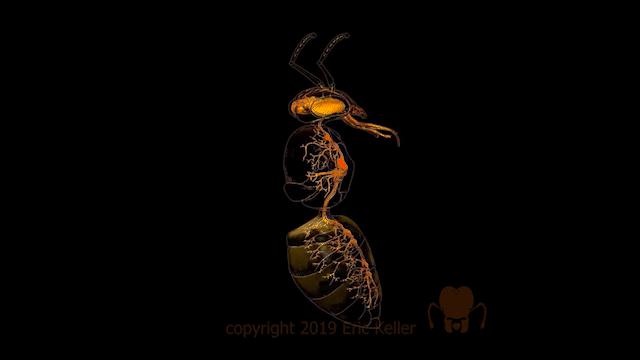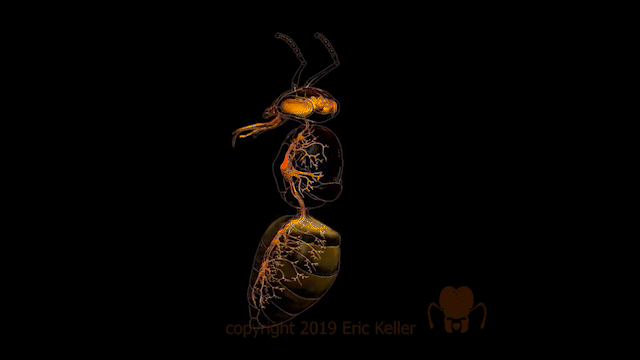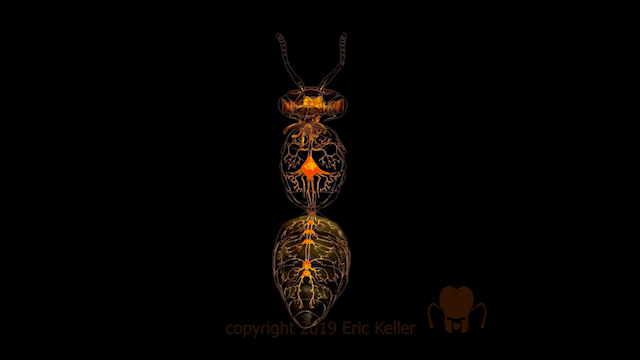
Le système nerveux des insectes
Nombre de vues : 12 358 Par Benoît GILLES Description générale Identique à celui des autres arthropodes, le système nerveux des insectes, est constitué d’une chaîne
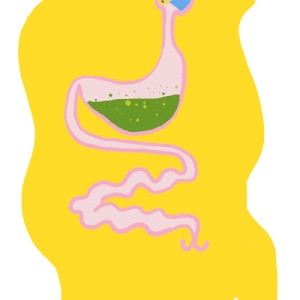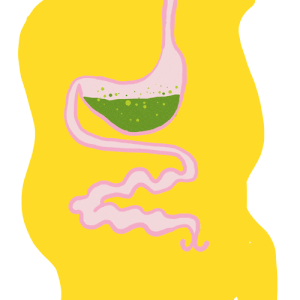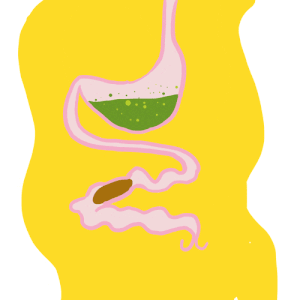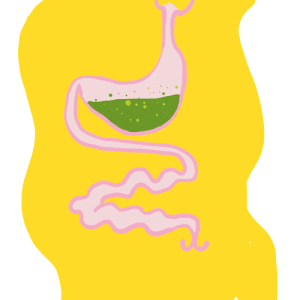
Le tube digestif des insecte – Anatomie
Par Benoît GILLES Le tube digestif des insectes est subdivisé en trois parties (figure 1) : Intestin antérieur (stomodeum) d’origine ectodermique Intestin moyen (mesenteron) d’origine endodermique Intestin postérieur (proctodeum) également d’origine ectodermique Contrairement au tube digestif des vertébrés couvert d’une paroi ayant de nombreuses couches (muqueuse, sous-muqueuse, musculeuse et tunique externe), celui des insectes est constitué…

Champignons entomopathogènes : Synthèse
Par Benoît GILLES Avec près de 100 000 espèces décrites, le règne des champignons constitue l’un des groupes les plus diversifiés des eucaryotes (organismes cellulaires à noyaux) et ayant conquis l’ensemble des écosystèmes terrestres et aquatiques. Ce nombre représente une infime fraction de la diversité connue estimée, quant à elle, entre 1,5 millions et 5…
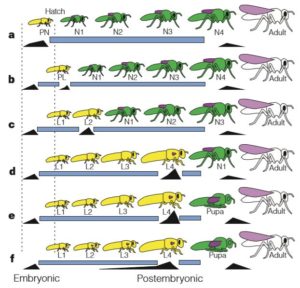
Origine de la métamorphose chez les insectes
Par Benoît GILLES La métamorphose est l’une des stratégies de vie les plus répandues chez les animaux. Les fortes différences morphologiques et physiologiques entre les formes larvaires et adultes impliquent l’exploitation d’habitats et de ressources alimentaires différentes, induisant des adaptations extrêmes pour des fonctions spécifiques telle que la dispersion. Chez les amphibiens et de nombreux…