
Par Benoît GILLES
Pour qu’ils se multiplient, la nature a doté les organismes vivants d’une large diversité de systèmes reproductifs. Chez les insectes, l’une de ces stratégies est la parthénogenèse. Celle-ci repose sur le développement d’individus issus de gamètes non fertilisés, donc sans recours à la fécondation. Cette stratégie est intéressante à plus d’un titre : en témoigne la déclinaison d’une multitude de formes (thélytoquie, arrhénotoquie et deutérotoquie) et son apparition à de multiples reprises au cours de l’évolution, au sein de taxons et d’espèces non apparentés et phylogénétiquement éloignés.
Description des différents types de parthénogenèse
I) Thélytoquie
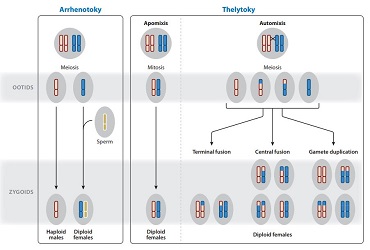
Correspondant à la parthénogenèse sensu stricto, la thélytoquie se caractérise par le fait que l’ensemble des œufs non fécondés émis par un insecte femelle n’engendre à leur tour qu’une descendance femelle diploïde.
La parthénogenèse thélytoque peut résulter deux systèmes génétiques distincts : l’un reposant sur la mitose (apomixie) et l’autre sur la méiose (automixie).
- L’apomixie est le système le plus simple : la descendance femelle est génétiquement similaire à la mère, l’absence de méiose évite tout réarrangement chromosomique (brassage génétique). Mère et fille peuvent ainsi être considérées comme des clones
- L’automixie fait intervenir le processus de méiose, la diploïdie est rétablie sans l’apport d’un gamète (sans fécondation). Ce processus peut être réalisé de plusieurs manières : 1) duplication du génome avant la méiose, créant une cellule avec 4n chromosomes (4 jeux de chromosomes) ; 2) fusion des deux noyaux durant la méiose ; 3) un noyau haploïde issu de la méiose se duplique par mitose pour fusionner à nouveau avec lui-même
II) Arrhénotoquie
Dans une parthénogenèse arrhénotoque, la descendance provenant d’œufs non fécondés n’est constituée que d’individus mâles (système opposé à la thélytoquie). Il existe deux systèmes génétiques distincts : 1) haplodiploïdique, les mâles sont haploïdes (1 jeu de chromosomes) et les femelles diploïdes (2 jeux de chromosomes) ; 2) diploïdique, mâles et femelles sont issus d’œufs diploïdes comme pour une parthénogenèse thélytoque.
III) Deuthérotoquie
Une parthénogenèse deuthérotoque engendre des œufs non fécondés à la fois des mâles et de femelles.
Apparition de la parthénogenèse
Un système parthénogénétique peut apparaître de diverses manières. Le cas le plus singulier repose sur l’interaction entre certains groupes d’insectes et des bactéries endosymbiotiques telles que Wolbachia, Rickettsia et Cardinium – la plus largement représentée étant Wolbachia pipientis – causant des incompatibilités cytoplasmiques provoquant soit l’arrêt du développement des embryons diploïdes, soit de la thélytoquie, soit de la féminisation des mâles, leur mort. Ces bactéries jouent souvent un rôle dans la conversion des sexes et modifie le sex-ratio en faveur des femelles.
De nombreuses espèces d’insectes sont monosexuées (leur population n’est composée que d’un seul sexe). Ces espèces se rencontrent dans des environnements isolés comme des îles ou des régions de haute altitude. Elles réalisent une parthénogenèse géographique, démontrant le pouvoir adaptatif de ce type de reproduction.
Parthénogenèse chez les hémimétaboles (ou hétérométaboles)
Les insectes hémimétaboles fédèrent les espèces dont la métamorphose est dite incomplète : le stade adulte est atteint progressivement au cours du cycle de développement par mues successives, les larves possédant la plupart des attributs des adultes, hormis les organes sexuels et parasexuels (lien article).
Odonata (libellules) : seule l’espèce Ischnura hastata (Caenagrionidae), se rencontrant aux Azores, a été décrite comme parthénogénétique (type thélytoque).
Orthoptera (criquets, grillons et sauterelles) : les espèces Locusta migratoria et Schistocerca gregaria ont la capacité d’engendrer de manière spontanée de la descendance femelle provenant d’œufs non fécondés : thycoparthénogenèse. Les gamètes haploïdes deviennent diploïdes progressivement au cours du développement. Pour Loxoblemmus frontalis, seule espèce de la famille des Gryllidae à pratiquer la parthénogenèse, la thélytoquie est induite par la présence de bactéries Wolbachia. Elle l’est sans intervention de Wolbachia chez la sauterelle Saga pedo (Tettigonidae).

Phasmatodea (Phasmes) : la parthénogenèse est plutôt bien répandue. Par exemple, le genre Timema, endémique de Californie, est composé de 5 espèces toutes apparentées et descendantes d’une même lignée. De rares mâles fertiles ont cependant pu être collectés, leur présence étant estimée à moins de 0,2% de la population. La détermination sexuelle des phasmes repose sur un système où les individus porteurs d’une paire de chromosomes autosomes (XX) sont femelles et ceux n’ayant qu’un exemplaire du X sont mâles (XO), (chez l’humain le système est de type XX-XY), les mâles se développent via la perte spontanée du chromosome X durant l’oogenèse. D’autres espèces se reproduisent également uniquement par parthénogenèse comme Bacillus rossius et Clonopsis gallica (espèce se rencontrant dans le sud de la France) (lien : histoire évolutive des Phasmatodea).
Blattoptera (anciennement Isoptera) (termites) : plusieurs groupes ont recours de manière facultative à de la parthénogenèse thélytoque par un processus de restauration de la ploïdie similaire à celui présent chez Reticulitermes speratus et R. virginicus (automiscie). Ce genre de termites est connu pour abriter des Wolbachia, que les colonies soient parthénogénétiques ou bisexuées, suggérant l’absence de corrélation entre les deux modes de reproduction.
Mantodea (mantes) (lien) : seules deux espèces, Miomantis savignyi et Bruneria borealis, procèdent à la parthénogenèse (thélytoque), parthénogenèse obligatoire chez B. borealis.
Hemiptera (cigales, pucerons et punaises) : ce ordre offre quant à lui une abondante diversité de systèmes parthénogénétiques au sein d’un grand nombre de familles comme Aclerdidae, Diaspididae, Aphididae ou encore Anthocoridae. Par exemple, plusieurs espèces de Fulgores (Delphacidae) appartenant aux genres Delphacodes et Ribautodelphax ont recours à la thélytoquie et à la pseudogamie (les femelles s’accouplent avec des mâles mais la descendance est totalement constituée de femelles). L’intervention des Wolbachia dans le processus de parthénogenèse a été démontrée chez Delphacodes kuscheli mais pas chez d’autres espèces. Une multitude de formes de parthénogenèses se rencontre entre les espèces des familles Coccidae et les Diapsididae : arrhénotoquie avec les mâles pouvant être diploïdes ou haploïdes, et deutérotoquie.
Certaines espèces de pucerons (Aphidimorpha) présentent un mode reproductif cyclique, passant d’un mode bisexué à parthénogénétique selon les saisons. Ainsi, au printemps, une femelle puceron (fondatrice) se multiplie par parthénogenèse afin de coloniser rapidement le milieu, puis, à l’automne, la reproduction devient sexuée aboutissant à la production d’œufs fécondés qui passent l’hiver à l’abri dans la végétation.
Concernant les punaises « véritable », il n’y pas de cas révélé de parthénogenèse. Cependant, deux espèces ont recours de manière facultative à la télytoquie : Calliodis maculipennis (espèce néotropicale de la famille des Anthociridae) et Campyloneura virgula (Miridae). Chose surprenante, il a été découvert une population parthénogénétique de C. maculipennis située sur l’île de Trinidad alors que les populations du continent, du Mexique et de Guyane, sont sexuées.
Cycle biologique du puceron (en anglais)
Parthénogenèse chez les holométaboles
Le groupe des holométaboles caractérise les espèces ayant recours à une phase de métamorphose dite complète où le passage à l’état adulte requière une phase chrysalide (papillon) ou pupe (mouches) durant laquelle la larve se transforme totalement (lien article).
Les holométaboles composent la très grande majorité de la diversité des espèces d’insectes : 800 000 espèces réparties en 11 ordres comme celui des Diptères, des Coléoptères, des Lépidoptères ou encore des Hyménoptères.
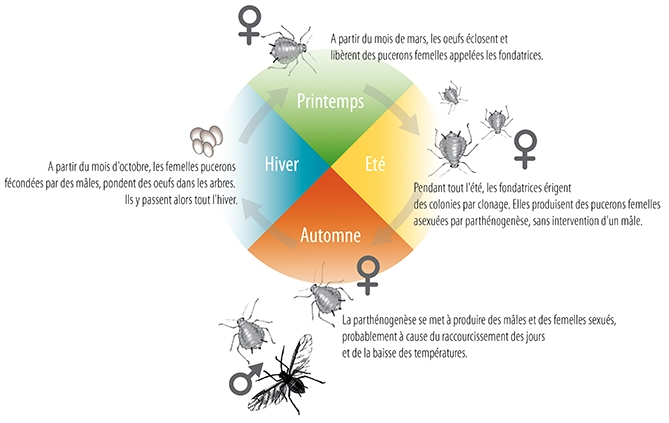
Les Hyménoptères (guêpes, abeilles, fourmis), avec près de 150 000 espèces décrites, constitue l’un des ordres d’insectes les plus diversifiés. La parthénogenèse est commune à toutes les espèces de l’ordre, regroupant ainsi la plus grande diversité de types de parthénogenèse. Le plus répandu et ancestral est l’arrhénotoquie combinée avec de l’haplodiploïdie. Les femelles ont ainsi la capacité de fertiliser ou non leurs œufs et pouvent ajuster le sex-ratio de leur descendance : un œuf diploïde donnera une femelle ou une ouvrière stérile et un œuf haploïde un mâle (voir illustration ci-dessous). Les mécanismes peuvent cependant varier entres les taxons.
Le passage d’une parthénogenèse arrhénotoque à une parthénogenèse thélytoque est relativement fréquent en raison de l’absence de chromosome sexuel rendant possible la restauration de la diploïdie par un processus apomictique ou automictique. Par exemple, l’espèce Diplolepis eglanteria (Cynipidae) est une petite guêpe ayant recourt à l’apomixie, et Apis mellifera capensis a, quant à elle, recourt à l’automixie.

La thélytoquie et l’arrhénotoquie peuvent également s’observer de manière cyclique, les ouvrières des fourmis Cataglyphis hispanica, Paratrechina longicornis, Vollenhovia emeyri et Wasmannia auropunctata sont issues de la reproduction sexuée alors que les nouvelles reines proviennent d’œufs parthénogénétiques thélytoques. L’étude du mode de reproduction de Wasmannia auropunctata a mis en évidence un cas unique de double système parthénogénétique (arrhénotoque et thélytoque). Il a ainsi été découvert en 2005 que les mâles, issus d’œufs fécondés, exprimaient uniquement le génome paternel, le génome maternel disparaissant (hormis le génome mitochondrial) par un mécanisme demeurant en partie inconnu. Ce processus indique que la descendance mâle est réalisée par clonage (voir illustration ci-contre).
Dans la mesure où ces ouvrières sont stériles, cette reproduction sexuée n’aboutit pas au mélange des génomes mâles et femelles à la génération suivante. Cette absence de flux de gènes entre mâles et femelles conduit à une différenciation génétique et à une évolution séparée des deux génomes. Cela pose la question quant à la classification d’une éventuelle distinction entre ces deux sexes comme deux espèces distinctes, dont l’une ne serait constituée que de mâles ! De plus, les mâles peuvent être considérés comme des parasites exploitant les femelles, où la production d’ouvrières stériles assure la protection et le ravitaillement de la colonie.
Chez 30% des espèces de guêpes parasitoïdes (lien articles) Cynipidae et Chalcidoidea, la thélytoquie est provoquée par des micro-organismes endosymbiotiques du genre Wolbachia, Cardinium et Rickettsia.
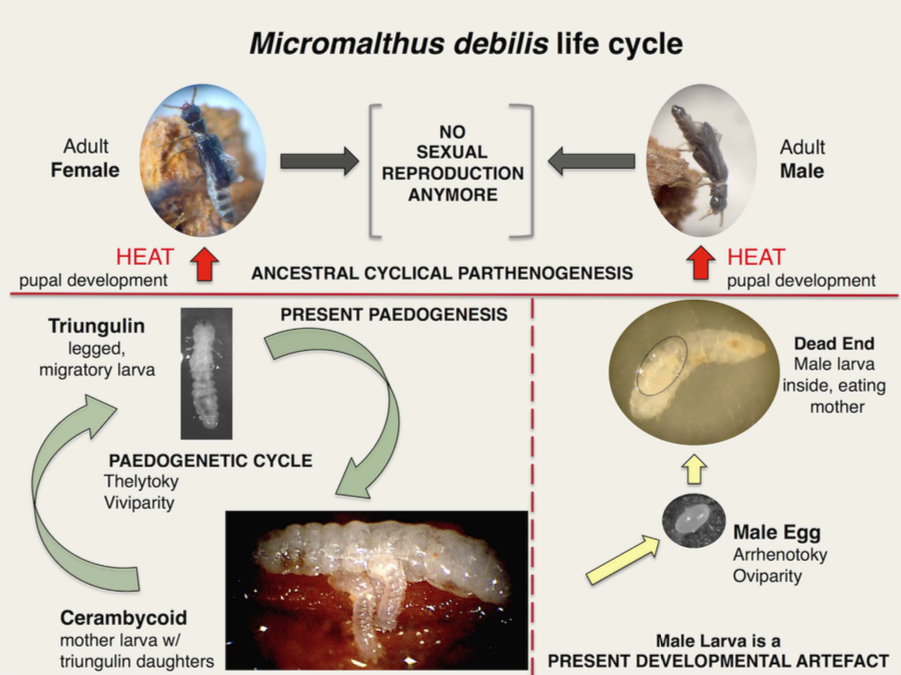
L’ordre des Coléoptères compte 30% des espèces d’insectes, soit 380 000 espèces dont seulement 600 espèces (20 familles) sont parthénogénétiques. La thélytoquie est plus répandue que l’arrhénotoquie : elle se rencontre entre autres chez les Alexiidae, Anobiidae, Cerambycidae, Dermestidae, Elateridae, Hydrophiliidae, Passalidae, Sphindidae ou encore Staphylinidae. Le cas de Micromalthus debilis est unique en son genre, la thélytoquie est poussée à l’extrême car les mâles, stériles, ont quasiment disparu des populations (lire cet article).
Ainsi, l’espèce Reesa vesopulae (Dermestidae), présente sur l’ensemble de la région néarctique, est strictement parthénogénétique, comme la majorité des populations nord-américaines de Cis fuscipes (Ciidae) et de Aelus mellillus (Elateridae). Chez les Chrysomelidae, l’espèce Bromius obscurus est représentée par des populations bisexuelles diploïdes en Amérique du Nord et d’autres triploïdes apomictiques en Europe. Les deux espèces européennes de charançons (Curculionidae), Polydrusus mollis et Otiorhynchus scaber sont principalement thélytoques, quelques populations sont cependant bisexuées diploïdes dans de petites localités.
Chez les Lépidoptères, malgré plus de 170 000 espèces décrites, seules deux douzaines d’espèces pratiquent la parthénogenèse. Ces espèces font parties principalement des Lymantriidae et des Psychidae (11 espèces). Il existe une forme de Dahlica triquetrella sexuée en Europe Centrale, alors que plusieurs populations thélytoques de type diploïde et tetraploïde sont répandues à travers l’Europe et l’Amérique du Nord.
Concernant les mouches (Ordres des Diptères), la parthénogenèse est apparue au sein d’au moins 11 familles regroupant plus de 150 000 espèces : Chironomidae, Hybotidae, Agromyzidae, Cecidomyiidae, Psychodidae, Sciaridae, Ctenostylidae, Lonchopteridae, Simuliidae, Ceratopogonidae et Chamaemyliidae. La moitié des taxons de Chironomidae étudiée apparaît parthénogénétique. Cette stratégie semble avoir été sélectionnée pour survivre à des conditions environnementales extrêmes, froid notamment (altitude et latitude), comme Eretmoptera murphyi (Antarctique) et Micropsectra sedna (Canada). D’autres, comme Cladotanytarsus aeiparthenus et Paratanytarsus grimmii, vivent dans des eaux polluées ou acides.
La parthénogenèse peut également survenir chez des espèces évoluant dans des habitats isolés ou fragmentés comme des grottes pour Troglocladius hajdi et Lymnophyes minimus vivant dans les îles Gough et Nightingale au sud de l’Océan Atlantique, ou Monopelopia caraguata, Phtytelmatocladius delarosai et Polypedilum parthenogeneticum qui vivent quant à elles dans de petites flaques (eau accumulée à l’aisselle de feuille, cavité de tronc, etc.) situées sur des plantes terrestres : phytotelme, du grec ancien phyto – plante ; telma – mare.
Tres interessant et tres bien ecrit. Merci.