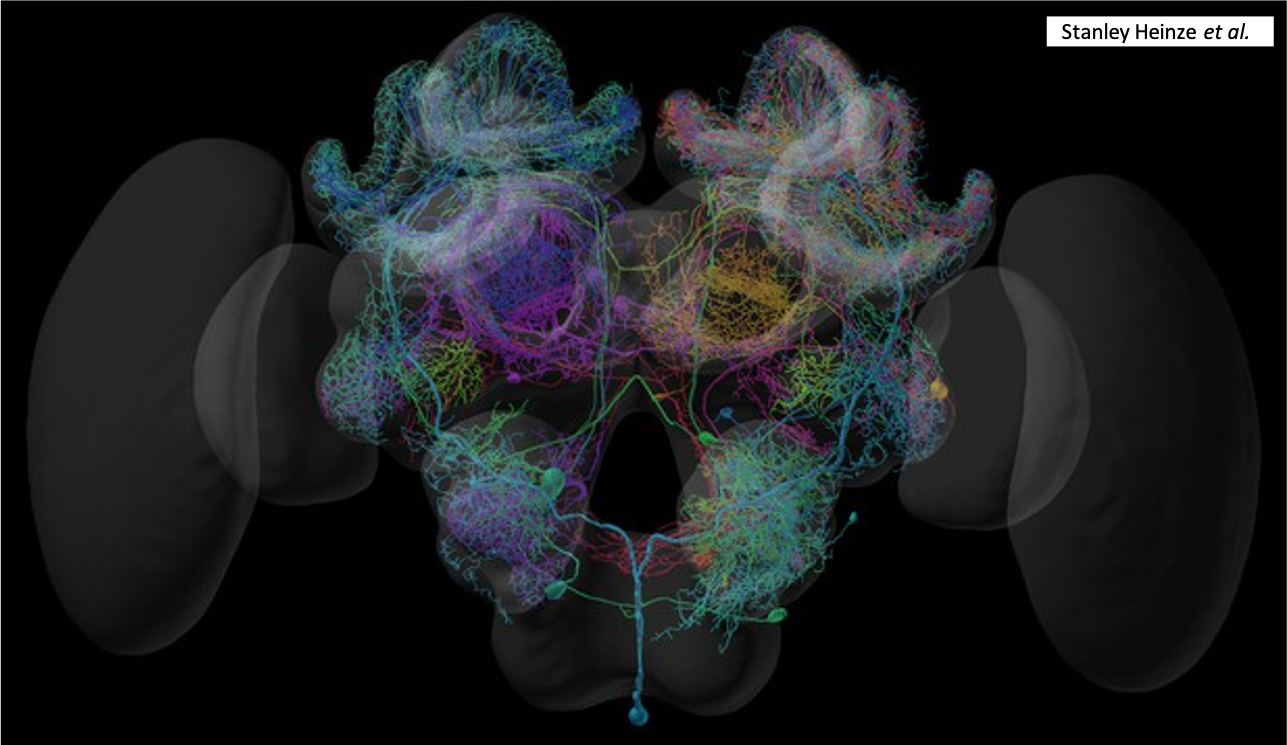
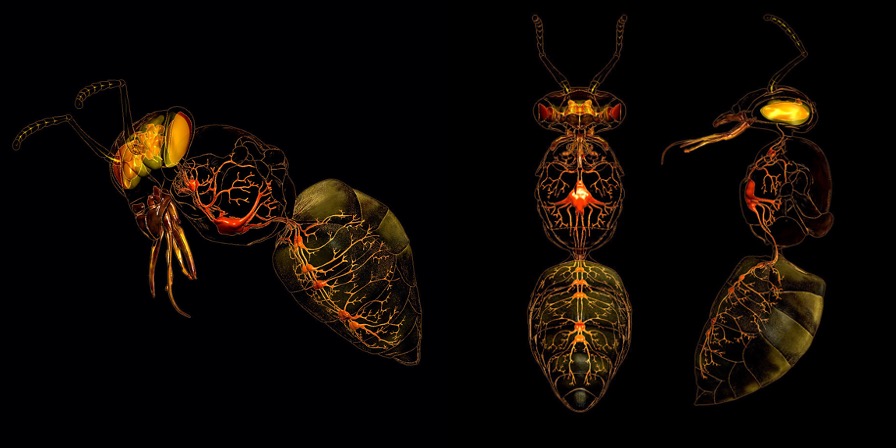
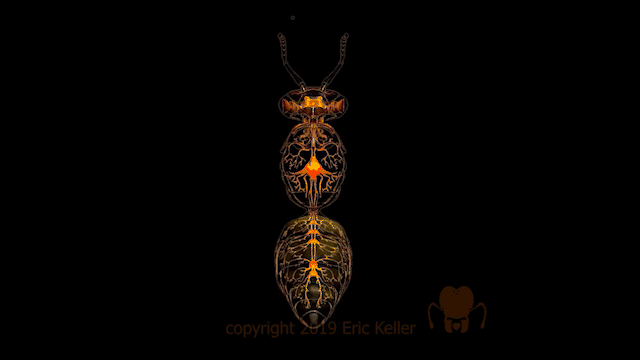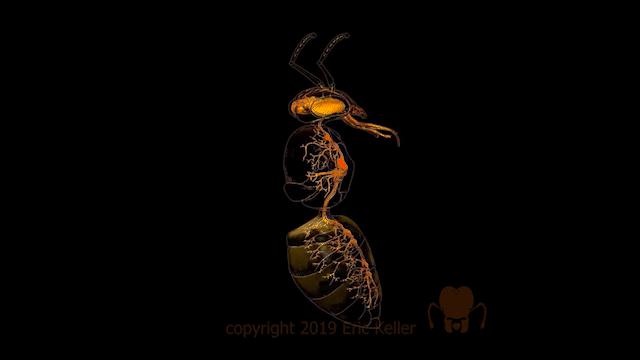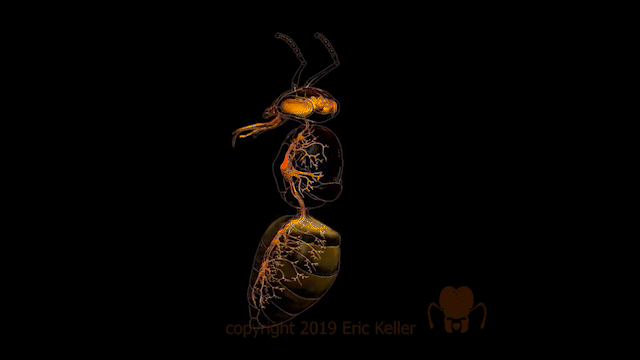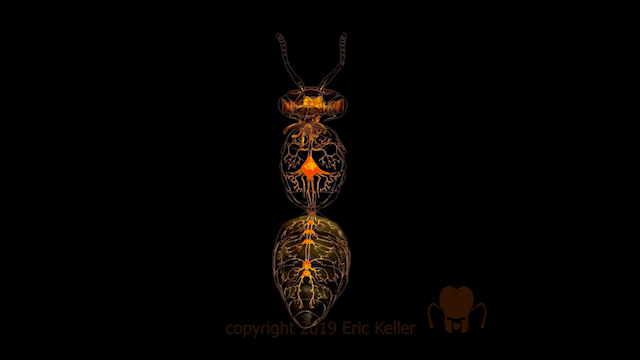
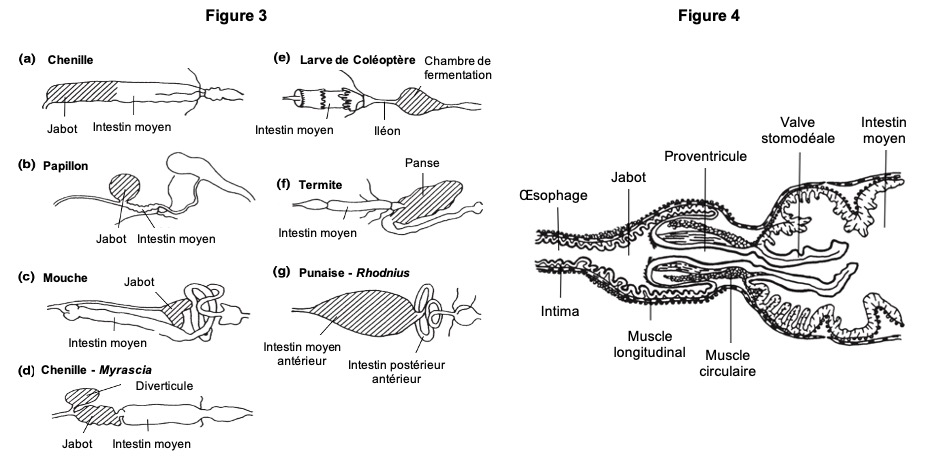
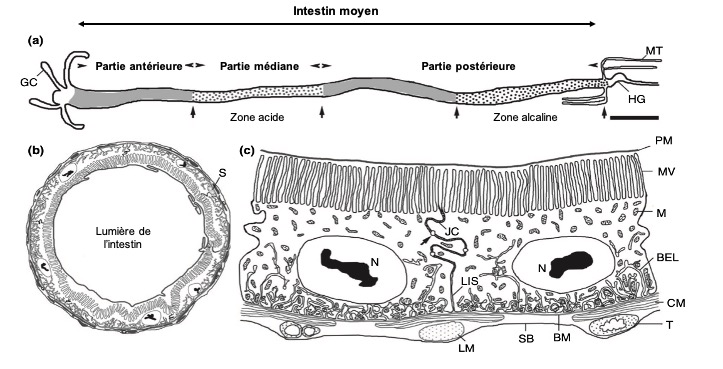
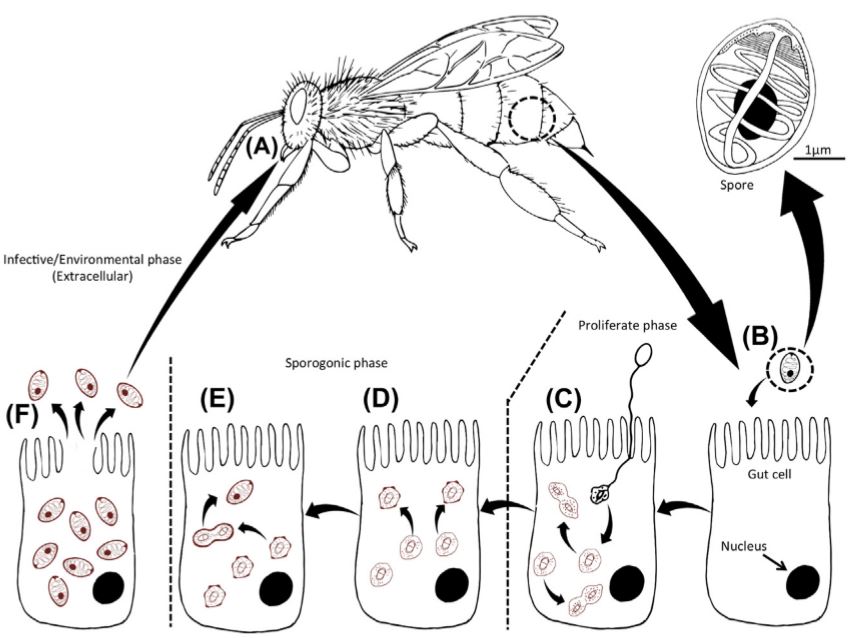
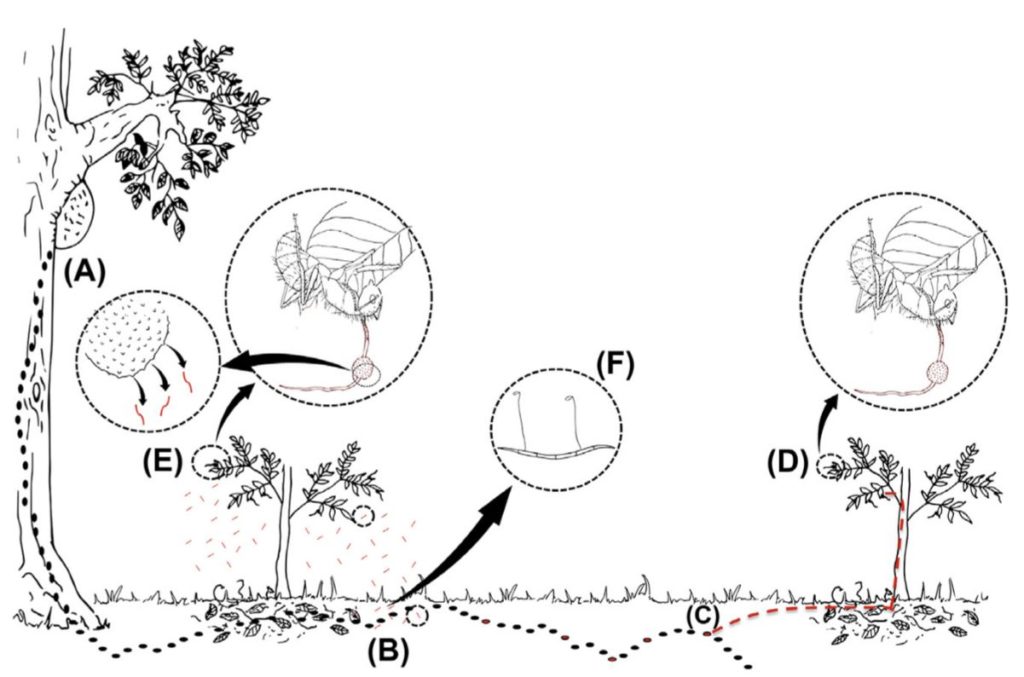
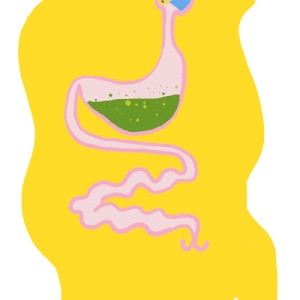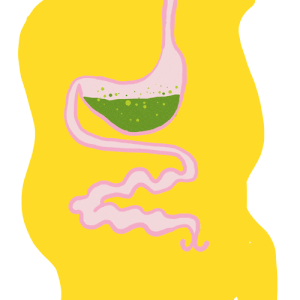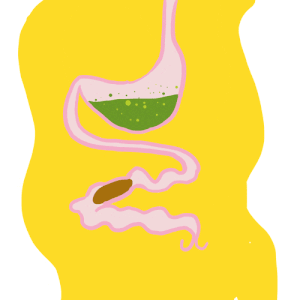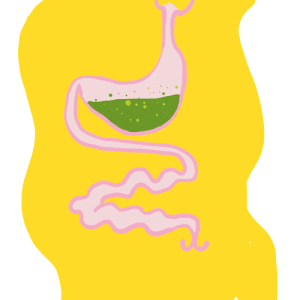Chez les insectes, le système circulatoire se distingue de celui des vertébrés à la fois par sa morphologie et par son fonctionnement. Il assure le transport de l’hémolymphe (liquide circulatoire chez les arthropodes) à travers un réseau relativement simple, composé principalement d’un vaisseau dorsal linéaire antéro-postérieur situé juste sous l’exosquelette (figures 1 – 2 – 4 – 8 – video en bas de page).
Figure 1 : Description détaillé du système circulatoire d’un insecte (Source : Chapman – Structure and Fonction, 2013 – modifié par B. GILLES)
Dans certains cas particuliers, comme chez les lépidoptères et certains hyménoptères, ce vaisseau s’enroule entre les muscles longitudinaux des ailes.
Figure 2 : Schéma général simplifié du système circulatoire chez l’insecte (Source : Quizlet – modifié par B. GILLES)
Il peut être soit fixé à la paroi dorsale du corps, soit suspendu à celle-ci par des filaments élastiques. À l’avant du corps, le vaisseau dorsal s’associe au tube digestif pour s’insérer entre le cerveau et l’œsophage. S’il est ouvert à l’avant, il est généralement fermé à l’arrière, à l’exception des larves d’éphémères (Éphéméroptères), chez lesquelles trois vaisseaux se ramifient vers les filaments caudaux depuis l’extrémité du cœur.
Ce vaisseau dorsal se divise en deux parties distinctes :
Un cœur postérieur, dont la paroi est perforée par des ostiums, des orifices d’expulsion ou d’aspiration (figures 1 – 2 – 4)
Une aorte antérieure, qui prolonge le cœur sous la forme d’un tube non perforé
Le cœur est généralement confiné à l’abdomen, mais chez les blattes (Blattodea), il peut s’étendre jusqu’au prothorax. Chez les orthoptères, il présente une structure segmentée en raison de dilatations localisées en ampoules (canal dilaté) au niveau des ostiums (orifice ou court canal ouvrant l’accès à une cavité ou faisant communiquer deux cavités). Chez certaines larves, comme celles des libellules (Odonates) et des tipules (Tipula), il est compartimenté en plusieurs chambres par des valves positionnées en amont de chaque paire d’ostiums d’aspiration. Enfin, chez les larves de Cloeon (Éphéméroptères), les valves ostiales sont si allongées qu’elles se rejoignent, formant une paroi (septum) au centre du vaisseau.
La paroi du vaisseau dorsal, contractile, est généralement constituée d’une ou de deux couches de cellules musculaires disposées en cercle ou en spirale (figure 3). Des fibres musculaires longitudinales sont également présentes ; chez les Hétéroptères, elles s’insèrent dans la paroi du vaisseau en avant et en arrière et ne sont pas connectées aux autres tissus.
Figure 3 : Développement et structure du vaisseau dorsal – (a) Étapes consécutives de la morphogenèse, de haut en bas. Les cardioblastes se polarisent et étendent leurs prolongements vers la ligne médiane dorsale jusqu’à ce qu’ils rencontrent leurs homologues controlatéraux. Les bords arrière des cardioblastes s’étendent ensuite postérieurement et médialement et entrent en contact, formant ainsi la lumière du vaisseau dorsal – (b) Le vaisseau dorsal mature est constitué de deux rangées de cardiomyocytes semi-circulaires. Leur décalage et leurs points de contact sur les lignes médianes dorsale et ventrale forment une disposition en spirale. La coupe antérieure montre l’organisation des cellules ostiales ; les deux lambeaux dirigés vers l’intérieur agissent ensemble comme une valve qui permet à l’hémolymphe de pénétrer dans le vaisseau, mais entrave son écoulement vers l’extérieur (Source : Hillyer & Pass, 2020)
Le muscle cardiaque des insectes peut adopter différentes orientations, bien que cette diversité apparente soit parfois liée à l’insertion des muscles alaires sur le cœur (figure 1). Les muscles du vaisseau dorsal se caractérisent par des sarcomères courts (dispositif musculaire générateur de force ou porteur de charge), avec des bandes d’environ 2 µm de long. Chaque filament épais est entouré de 9 à 12 filaments fins, une organisation similaire à celle des muscles viscéraux.
Figure 4 : Différents types de systèmes circulatoires et modes de flux chez les insectes. Flèches vertes indiquent l’afflux dans le vaisseau dorsal, et flèches bleues et rouges indiquent respectivement le flux intracardiaque antérograde et rétrograde – (a) Campodea augens (Diplura) représente une forme ancestrale, avec des artères antennaires et cercales, un anneau vasculaire circumœsophagien dans la tête et un flux bidirectionnel dans le vaisseau dorsal – (b) Schistocerca shoshone (Orthoptères) représente la condition de nombreux basalptérygotes, avec un flux antérograde dans les vaisseaux dorsaux et de nombreux ostiums appariés. Dans le mésothorax et le métathorax, le vaisseau dorsal présente des élargissements ampullaires qui aspirent l’hémolymphe des veines alaires par l’intermédiaire de bras scutellaires formés de cuticule tergale – (c) Drosophila melanogaster (Diptères) représente la condition dérivée avec des inversions du rythme cardiaque, et ainsi, le vaisseau dorsal alterne périodiquement entre contraction antérograde (en haut) et rétrograde (en bas). L’hémolymphe est irriguée par des organes pulsatiles distincts aux antennes et aux ailes. Abréviations : aa, artères antennaires ; ah, cœur antennaire ; cv, vaisseau cercal ; cvr, anneau vasculaire circumœsophagien avec ouverture ventrale ; eo, ostium à flux sortant ; io, ostium à flux entrant ; iv, valve intracardiaque ; sa, bras scutellaire ; te, élargissement thoracique du vaisseau dorsal ; wh, cœur alaire (Source : Hillyer & Pass, 2020)
Ces muscles sont enveloppés, à l’intérieur comme à l’extérieur, par une lame basale qui recouvre également les néphrocytes (cellules ayant un rôle dans l’excrétion). Les ostiums à flux entrants (afférent), qui permettent l’entrée de l’hémolymphe dans le cœur, sont des ouvertures en forme de fentes situées sur la paroi latérale du cœur. Un insecte peut en posséder jusqu’à 12 paires, réparties en neuf paires abdominales et trois paires thoraciques (figure 4).
Cette répartition varie selon les ordres :
Blattodea et Orthoptera : 12 paires complètes
Lépidoptères : généralement sept ou huit paires
Hyménoptères aculéates (abeilles, guêpes, fourmis) : cinq paires
Mouche (Musca) : seulement quatre paires
Poux (Phthiraptera) et certains hétéroptères : deux à trois paires seulement, avec un cœur restreint aux segments abdominaux postérieurs.
Les ostiums possèdent deux lèvres, une antérieure et une postérieure, qui se rétractent vers le cœur pour former une valvule. Ce mécanisme permet l’entrée de l’hémolymphe lors de la diastole (expansion du cœur) tout en empêchant son reflux vers l’extérieur durant la systole (contraction) (figures 3 – 5 – 6). Lors de la diastole, la pression de l’hémolymphe entrante force les lèvres à s’écarter. Une fois cette phase terminée, la pression interne les referme, les maintenant closes jusqu’à la fin de la systole.
Figure 5 : Différents types d’ostiums présents dans le vaisseau dorsal – (a) Ostiums à flux entrants à lèvres paires et valve intracardiaque – (b) Ostiums à flux entrants à lèvres allongées fonctionnant comme une valve de poche – (c) Ostiums bidirectionnels à une seule lèvre, (gauche) Flux entrant pendant la phase antérograde – (droite) Flux sortant pendant la phase rétrograde – (d) Ostiums à flux entrant (en bas), ostiums à flux sortant avec des valves sphinctériennes à la base des vaisseaux latéraux (au milieu), et un ostium à flux sortant impair (en haut). La valve sphinctérienne de gauche est représentée en position ouverte, tandis que celle de droite est représentée en position fermée (Source : Hillyer & Pass, 2020)
Chez certaines espèces, la structure de ces valvules varie :
Chaoborus (mouches fantômes) : les valvules ont tendance à s’évaginer sous la pression exercée pendant la systole, mais une paroi unicellulaire interne empêche leur éversion complète (figure 6 (a))
Bombyx mori (ver à soie) : seule la lèvre postérieure de chaque ostium se replie à l’intérieur du cœur (figure 2b). Lors de la systole, ce repli est plaqué contre la paroi cardiaque, empêchant toute fuite d’hémolymphe (figure 6 (b))
Figure 6 : Fonctionnement des ostiums à flux entrant – (a) Larve de Chaoborus ; les valves sont empêchées de s’ouvrir vers l’extérieur lors de la systole par un filament unicellulaire (non représenté) fixé à l’intérieur du cœur (d’après Wigglesworth, 1972) – (b) Larve de Bombyx ; le cœur est représenté en coupes horizontale (à gauche) et transversale (à droite) (Source : Chapman – Structure and Fonction, 2013 – modifié par B. GILLES)1972). (b) Larve de Bombyx ; le cœur est représenté en coupes horizontale (à gauche) et transversale (à droite) (Source : Chapman – Structure and Fonction, 2013 – modifié par B. GILLES)
Lorsque le rythme cardiaque s’inverse (voir video ci-dessous), l’hémolymphe s’écoule via les ostiums afférents. En revanche, les ostiums à flux sortants (efférent) sont plus rares et ne se retrouvent que chez certains groupes d’insectes, notamment chez les Thysanoures (poissons d’argent), Orthoptères, Plécoptères et Embioptères. Leur disposition varie selon les ordres :
Plécoptères et Embioptères : ostiums efférents impairs
Orthoptères : paires d’ostiums ventro-latéraux sans valves internes, généralement cinq paires abdominales et deux thoraciques
Blattodea et Mantodea : absence d’ostiums afférents ; l’hémolymphe s’évacue via des vaisseaux segmentaires latéraux
Periplaneta (blatte) : cinq paires de vaisseaux segmentaires abdominaux chez les nymphes tardives, avec deux paires thoraciques supplémentaires à l’âge adulte (voir video ci-dessous)
Mantes : uniquement des vaisseaux abdominaux, chacun équipé d’une valve musculaire unidirectionnelle
Les parois de ces vaisseaux segmentaires ne sont pas musculaires, ce qui limite ainsi leur rôle au simple transport de l’hémolymphe.
Figure 7 : Composés produits par les neurones agissant comme des neurotransmetteurs ou des neuromodulateurs. Certains peuvent également agir comme neurohormones (Source : Chapman – Structure and Fonction, 2013)
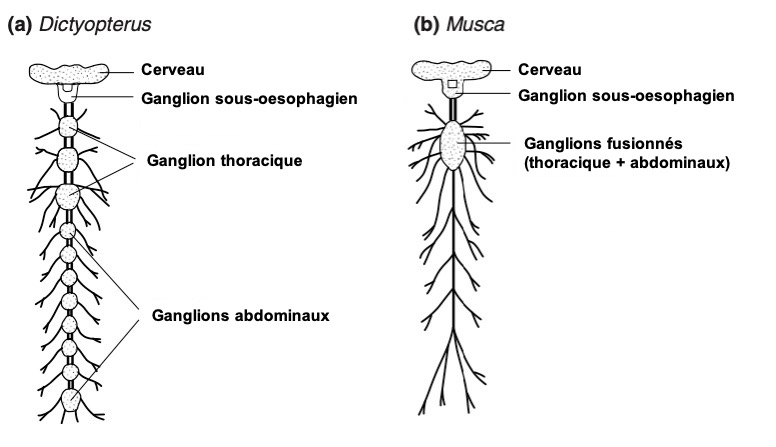
Chez la plupart des insectes, le cœur est innervé par des nerfs issus desganglions segmentaires ventraux, qui longent la paroi corporelle. Dans plusieurs ordres, comme chez lesOdonates, Blattodea, Phasmatodea, Orthoptères, leslarves de Lépidoptèreset certainsColéoptères adultes, ces nerfs segmentaires se regroupent pour former unnerf cardiaque latéral, qui court de chaque côté du cœur.
ChezLocusta, par exemple, des groupes de neurones situés sur la ligne médiane de chaque ganglion abdominal envoient des axones vers le cœur. De plus, unepaire de cellules neurosécrétricesdu ganglion sous-œsophagien projette un axone vers l’avant, à travers leconjonctif circumœsophagien, et un autre vers l’arrière, le long ducordon nerveux ventral. Ce dernier se ramifie à chaque ganglion abdominal et donne naissance à unnerf latéral, qui s’étend dorsalement jusqu’au cœur. Ces branches, intégrées aunerf cardiaque latéral, présentent desterminaisons nerveuses dilatées (variqueuses), caractéristiques des cellules neurosécrétrices, qui libèrent unpeptide de type FMRFamide(figure 7).
Les nerfs cardiaques latéraux reçoivent également l’innervation de neuronesextra-ganglionnaires, situés à proximité du cœur, appelésneurones cardiaques.Periplaneta(blatte) en possède environ32, dont certains sontneurosécréteurs.
Chez la majorité desinsectes holométaboles, lesnerfs segmentairesatteignent directement le cœur sans former de nerfs cardiaques latéraux. Cependant, certaines exceptions existent : chez lesmoustiques Anopheles, le cœur ne reçoitaucune innervation directe, bien que des nerfs segmentaires soient connectés aux muscles alaires (figure 8).
Figure 8 : Chez le moustique Anopheles gambiae – (a) Le cœur d’un moustique adulte alterne périodiquement entre contractions antérogrades (en haut) et rétrogrades (en bas) (Source : Hillyer & Pass, 2020 – Modifié par B. GILLES)
Sinus et diaphragmes
L’hémocèle (cavité interne où circule l’hémolymphe) de nombreux insectes est organisé en trois sinus principaux (figure 9) :
Le sinus péricardique dorsal, situé juste sous le dos de l’insecte
Le sinus périviscéral, occupant la majeure partie de la cavité corporelle
Le sinus périneural ventral, localisé sous le cordon nerveux
Ces sinus sont séparés par deux diaphragmes : le diaphragme dorsal, qui délimite le sinus péricardique, et le diaphragme ventral, qui sépare le sinus périneural du sinus périviscéral. Chez la plupart des insectes, le sinus périviscéral domine en volume, mais il est particulièrement élargi chez les Ichneumonidés.
Figure 9 : Sinus principaux en coupe schématique (Source : Chapman – Structure and Fonction, 2013 – modifié par B. GILLES)
Le diaphragme dorsal et les muscles alaires
Le diaphragme dorsal est une fine membrane conjonctive perforée, généralement incomplète sur ses parties latérales. Cette discontinuité permet au sinus péricardique, situé au-dessus, de rester en continuité avec le sinus périviscéral, situé en dessous. Les limites latérales du diaphragme sont délimitées par la présence de muscles, de trachées, ainsi que par l’insertion des muscles alaires, qui s’intègrent directement à cette structure.
Les muscles alaires, qui s’étendent transversalement sous le cœur, jouent un rôle dans la circulation de l’hémolymphe. Chez Hyalophora (un papillon), ces muscles sont directement connectés aux muscles cardiaques via des disques intercalaires. Ils prennent naissance sur le tergum (partie dorsale d’un segment), s’étalant en éventail pour se rejoindre en une large zone médiane (figure 9). Chez les sauterelles et certains Orthoptères, leur origine est plus large, et seule la partie proche de leur attache est contractile, le reste étant constitué de faisceaux conjonctifs qui se ramifient et s’anastomosent. Certaines de ces fibres forment un plexus reliant le diaphragme à la paroi cardiaque.
Le nombre de muscles alaires varie selon les ordres d’insectes :
Orthoptères : jusqu’à dix paires abdominales et deux paires thoraciques
Hétéroptères terrestres : entre 4 et 7 paires
Chez d’autres insectes, leur nombre est réduit
D’un point de vue structurel, les muscles alaires sont des muscles viscéraux, composés d’environ 10 à 12 filaments fins pour chaque filament épais.
Le diaphragme ventral et sa variabilité
Le diaphragme ventral est un septum horizontal (structure anatomique faite d’un tissu dense formant une cloison séparant deux cavités), situé juste au-dessus du cordon nerveux ventral, séparant le sinus périneural du sinus périviscéral (figure9). Sa présence varie selon les groupes d’insectes et leur stade de développement :
Il est présent chez les larves et adultes des Odonates, Blattodea, Orthoptères, Neuroptères (chrysopes) et Hyménoptères
Il apparaît uniquement chez les adultes chez les Mécoptères (mouches-scorpions), Lépidoptères et Diptères nématocères
Chez les Lépidoptères, le cordon nerveux est relié à la face ventrale du diaphragme par du tissu conjonctif
Généralement, ce diaphragme est limité à l’abdomen, mais chez certaines sauterelles et hyménoptères, il s’étend aussi jusqu’au thorax.
Sa structure varie selon les espèces :
Chez les sauterelles, il s’agit d’une membrane mince et peu musclée
Chez d’autres insectes, il peut être une membrane musculaire solide
Latéralement, il se relie aux sternites, formant de larges ouvertures qui assurent la continuité entre les sinus périviscéraux et périneuraux
Son organisation évolue également au cours du développement : chez les Corydales (Neuroptera), il est solide chez la larve, mais devient fenêtré chez l’adulte.
Organes pulsatiles accessoires
Les insectes possèdent plusieurs structures pulsatiles facilitant la circulation de l’hémolymphe, en particulier dans les appendices tels que les ailes, les pattes, les antennes et, chez les femelles, l’ovipositeur (figures 10 – 11 – 12)
Les organes pulsatils des ailes
Chez la plupart des insectes adultes, un organe pulsatile spécifique assure l’irrigation des ailes en hémolymphe. Il est généralement présent dans les deux segments alaires, sauf chez les Diptères et les Coléoptères Polyphaga, où il est limité au mésothorax.
Cet organe comprend :
Un réservoir hémolymphatique (ou espace de stockage) situé sous la partie postérieure du tergum (scutellum). Ce réservoir est isolé partiellement ou totalement du reste de l’hémocèle thoracique
Une connexion avec les veines postérieures des ailes via le cordon axillaire
Une pompe musculaire, formée par la paroi ventrale du réservoir, qui peut être une extension du cœur ou une structure indépendante (figure 10)
Figure 10 : Organes pulsatiles accessoires. Sur tous les schémas, les flèches indiquent le sens du flux sanguin. (a)–(c) Cœur de l’aile non relié à l’aorte, comme chez la plupart des insectes holométaboles – (a) Coupe transversale du thorax montrant la connexion du réservoir sous-scutellaire au cordon axillaire de l’aile – (b), (c) Coupe longitudinale schématique, d’avant en arrière à gauche. Lorsque les muscles du diaphragme se contractent (b), celui-ci s’aplatit et le sang est aspiré par l’aile. Lorsque les muscles se relâchent (c), des éléments élastiques de suspension tirent le diaphragme vers le haut et expulsent le sang vers l’avant (d’après Krenn et Pass, 1993) – (d) Cœur de l’aile relié à l’aorte, comme chez la plupart des insectes hémimétaboles. Coupe longitudinale du thorax. Le réservoir sous-scutellaire est relié au cordon axillaire de l’aile de chaque côté (modifié d’après Whedon, 1938) (Source : Chapman – Structure and Fonction, 2013 – modifié par B. GILLES)
Mécanisme de pulsation
Le fonctionnement de cet organe pulsatile varie selon les groupes d’insectes :
Chez les insectes holométaboles et les Hémiptères
Un diaphragme musculaire indépendant du cœur entoure le réservoir sous-scutellaire sur sa face ventrale, relié au scutellum par plusieurs filaments élastiques (figure10a-c). La contraction des muscles (commandée par la moelle nerveuse ventrale) aplatit le diaphragme, aspirant l’hémolymphe des ailes vers le réservoir.
Lors de la relaxation musculaire, le diaphragme remonte, probablement sous l’effet de l’élasticité des filaments, refoulant l’hémolymphe dans la cavité corporelle (figure10b-c).
Chez les insectes hémimétaboles (hors Hémiptères), les Coléoptères et les Hyménoptères symphytes
L’organe pulsatile alaire est une expansion du vaisseau dorsal, formant un diverticule. Il possède une paire d’ostiums afférents s’ouvrant depuis le réservoir sous-scutellaire.
Chez les Odonates
Chaque segment du ptérothorax contient une ampoule pulsatile, reliée à l’aorte par un vaisseau étroit (figure 10d). Cette ampoule est suspendue au tergum par des ligaments élastiques et possède une paroi dorsale musclée. Lors de la contraction musculaire, l’ampoule est comprimée, refoulant l’hémolymphe dans l’aorte.
Simultanément, le volume du réservoir sous-scutellaire augmente, ce qui aspire l’hémolymphe depuis les ailes :
Les veines postérieures assurent le flux afférent (vers l’ampoule)
Les veines antérieures assurent le flux efférent (depuis l’ampoule vers la circulation générale)
Lors de la relaxation musculaire, les ligaments élastiques restaurent la forme de l’ampoule, permettant ainsi l’aspiration de l’hémolymphe depuis le réservoir
Figure 11 : Organes du segment abdominal 9 (à gauche) et du segment 8 (à droite) en deux phases d’action distinctes. Les muscles compresseurs ipsilatéraux des deux segments abdominaux (cm8, cm9) se contractent simultanément, en alternance avec l’autre côté. Ils compriment ainsi la chambre de pompage correspondante, c’est-à-dire la systole des organes, et l’hémolymphe est refoulée à contre-courant à travers les sinus des gonapophyses ovipositrices (ga) puis dans la cavité abdominale. Simultanément, les organes opposés sont en phase de diastole, durant laquelle les chambres de pompage se dilatent et se remplissent d’hémolymphe provenant de l’hémocèle abdominal. Les mouvements latéraux simultanés de la chambre génitale favorisent probablement l’aspiration de l’hémolymphe pendant la diastole et empêchent le reflux pendant la systole. D’autres abréviations (d, dorsal ; l, latéral ; m, médial ; v, ventral ; aiv, sclérite intervalvulaire antérieur ; cs, coxosternite ; opo, organe pulsatile de l’ovipositeur ; gc, chambre génitale ; si, sinus) (Source : Hustert et al., 2014 – modifié par B. GILLES)
Les organes pulsatiles antennaires et leurs mécanismes
Les insectes possèdent un organe pulsatile à la base de chaque antenne, permettant d’assurer la circulation de l’hémolymphe. Cet organe se présente sous la forme d’une ampoule, reliée à un fin tube qui s’étend presque jusqu’à l’extrémité de l’antenne.
Variations selon les groupes d’insectes
Thysanoures (poissons d’argent), Archéognathes (perce-oreilles) et certains Plécoptères : leurs ampoules sont dépourvues de muscles. Elles servent uniquement à diriger le flux d’hémolymphe depuis l’ouverture de l’aorte vers l’antenne
Chez la plupart des insectes : les ampoules sont munies de muscles dilatateurs. La compression de l’ampoule, qui propulse l’hémolymphe dans l’antenne, est assurée par l’activité de filaments élastiques situés de part et d’autre de sa paroi
Cas particulier des Dermaptères (perce-oreilles) : ce sont les seuls insectes à posséder des muscles compresseurs au niveau des ampoules
Chez la les blattes du genre Periplaneta : un muscle unique relie les deux ampoules antennaires. Lorsqu’il se contracte, il provoque la dilatation simultanée des deux ampoules, permettant ainsi l’entrée de l’hémolymphe par un ostium (figure 12e–g – vidéo ci-dessus). Ces contractions sont d’origine myogénique (capable de se contracter sans stimulation nerveuse), mais peuvent être modulées par des signaux nerveux, en raison de la présence de terminaisons nerveuses dans le muscle
La majorité des terminaisons nerveuses sont concentrées dans l’ampoule, qui pourrait ainsi jouer un rôle de centre neurohémal (organe ou aire dans lequel coexistent les nerfs et la circulation sanguine ou hémolymphatique). Les axones responsables de cette activité prennent naissance dans le ganglion sous-œsophagien et proviennent :
D’un neurone médian dorsal unique
D’une paire de somates situés latéralement
Figure 12 : Organes pulsatiles accessoires. Sur tous les schémas, les flèches indiquent le sens du flux sanguin. (e)–(g) Organe pulsatile antennaire de blatte – (e) Vue de dessus. Dessus de la tête coupé et cerveau retiré – (f) Le muscle dilatateur se contracte, élargissant la lumière de l’ampoule, permettant ainsi l’aspiration du sang depuis l’hémocèle de la tête. La baisse de pression provoque une constriction à l’origine du vaisseau antennaire, limitant ainsi le reflux sanguin depuis l’antenne – (g) Le muscle se relâche et l’ampoule s’aplatit grâce à l’élasticité de sa paroi interne et à la traction du tendon. L’ostium est fermé par une valve et le sang est injecté dans le vaisseau antennaire (d’après Pass, 1985) – (h) Organe pulsatile de la patte de Triatoma. La contraction du muscle comprime le sinus sanguin d’un côté du septum et l’élargit de l’autre, de sorte que le sang circule le long de la jambe d’un côté du septum et remonte de l’autre (d’après Kaufman et Davey, 1971) (Source : Chapman – Structure and Fonction, 2013 – modifié par B. GILLES)
L’aorte et les vaisseaux antennaires chez les Lépidoptères
Chez les Lépidoptères, l’aorte se termine antérieurement par un sac, à partir duquel émergent les vaisseaux antennaires, participant à la circulation de l’hémolymphe dans les antennes.
La circulation hémolymphatique dans les pattes
La plupart des insectes possèdent, au niveau des pattes, un septum longitudinal divisant leur lumière interne en deux sinus, permettant ainsi un flux bidirectionnel de l’hémolymphe.
Chez les Hémiptères : le septum subit une torsion de 90° à l’extrémité proximale du tibia. À cet endroit, on trouve un muscle spécifique (figure 12h) qui, lorsqu’il se contracte compresse un sinus, forçant l’hémolymphe à remonter vers le thorax. Simultanément, il élargit l’autre sinus, facilitant ainsi l’aspiration de l’hémolymphe dans la patte.
L’activité de ce muscle est probablement myogénique (capable de se contracter sans stimulation nerveuse), bien qu’elle puisse être modulée par le système nerveux.
Certains insectes, dotés de cerques particulièrement longs, nécessitent des adaptations spécifiques pour assurer la circulation de l’hémolymphe. Chez les éphéméroptères, de petits vaisseaux non contractiles s’étendent depuis l’extrémité postérieure du cœur jusqu’aux cerques, facilitant ainsi le flux hémolymphatique.
En revanche, chez les plécoptères, bien que des vaisseaux hémolymphatiques soient également présents dans les cerques, ils ne sont pas directement reliés au cœur. Ils s’ouvrent dans la cavité périviscérale, tandis que la lumière cercale reste connectée à cette cavité via le paraprocte (plaque ventro-latérale de l’extrémité de l’abdomen). Les variations de volume du paraprocte, générées par un petit muscle, permettent d’aspirer l’hémolymphe depuis la lumière externe du cerque et de la refouler dans la cavité périviscérale, assurant ainsi une circulation continue du fluide biologique.
Bibliographie
The Insects – Structure and Fonction – Cambridge University Press – Chapman – 2013 – 5ème édition
Hustert R. ; Frisch M. ; Böhm A. & Pass G. (2014) : A new kind of auxiliary heart in insect : functional morphology and neuronal control of the accessory pulsatile organs of the cricket ovipositor. Frontiers in Zoology, 11,43 (lien)
Hillyer J. & Pass G. (2020) : The insect circulatory system : structure, function and evolution. Annual Review of Entomology, 65:121-143 (lien)
Les sacs aériens sont une partie essentielle du système respiratoire des insectes, une innovation évolutive qui permet à de nombreuses espèces d’exploiter de nouvelles niches écologiques et d’optimiser des fonctions physiologiques comme le vol, la régulation de la flottabilité et la croissance. Ces structures dérivent du système trachéal, qui permet l’apport en oxygène et l’élimination du dioxyde de carbone, des gaz essentiels pour leur métabolisme énergétique. La structure et la fonction des sacs aériens varient grandement d’une espèce à l’autre, et leur développement est influencé par les besoins physiologiques et environnementaux.
Les premières observations des sacs aériens remontent au XVIIIe siècle, mais l’exploration systématique de leur rôle et de leur évolution a commencé avec des études sur les insectes volants tels que Drosophila melanogaster et d’autres arthropodes trachéates (respirant à l’aire libre vs aquatiques comme les crustacés) (Swammerdam, 1737 ; Guha et Kornberg, 2005).
Aujourd’hui, les sacs aériens sont considérés comme des innovations adaptatives dans la respiration des insectes, en particulier pour ceux nécessitant des capacités de vol élevées ou des adaptations spécifiques à leur environnement.
Phylogénie et apparition des sacs aériens
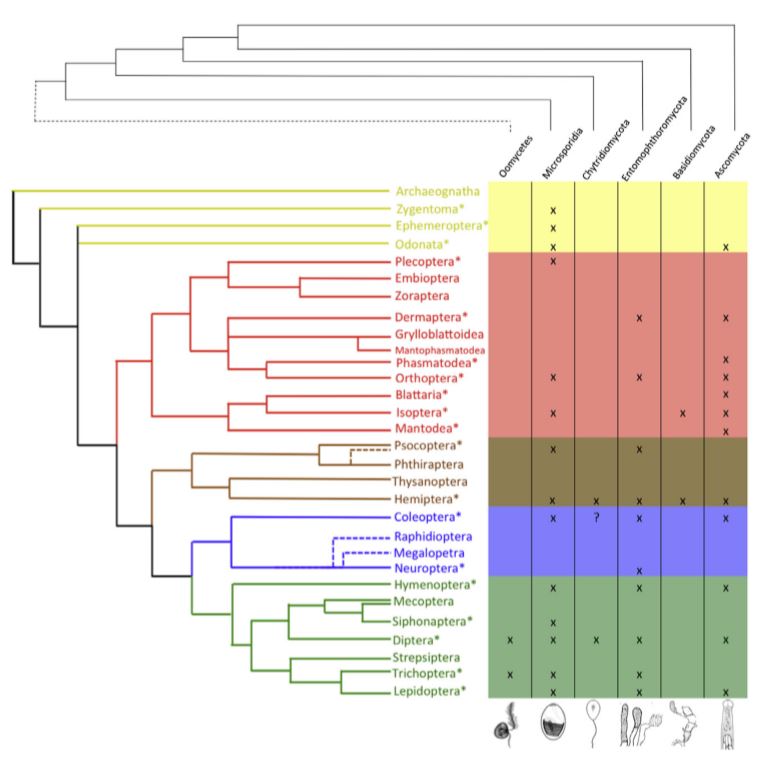
Chez les insectes primitifs, notamment les Paleoptera (Odonates et Éphéméroptères), les sacs aériens sont généralement rares ou limités dans leur développement. Les Odonates, des insectes capables de vol soutenu et puissant, font exception, avec des sacs aériens bien développés chez les adultes. En revanche, les Éphéméroptères, bien que volants, sont des « mauvais » volants et ne possèdent que des structures limitées ressemblant à des sacs aériens dans certains stades larvaires (Herhold et al., 2023).
La présence de sacs aériens est clairement associée aux Neoptera, un groupe d’insectes plus « évolués » qui inclut la majorité des ordres modernes. Parmi eux, les sacs aériens sont particulièrement fréquents dans les groupes ayant développé une capacité de vol élevée, comme les coléoptères, les diptères, les hyménoptères et les lépidoptères (figure 1).
Cette répartition phylogénique des sacs aériens et leur corrélation avec le vol suggèrent que ces organes sont apparus comme une adaptation essentielle pour améliorer l’efficacité respiratoire nécessaire au vol actif (Lee, 1929).
Figure 1 : Schémas phylogénétiques des sacs aériens et des systèmes trachéaux compressibles chez les Hexapodes adultes. Les sacs aériens sont associés à une forte capacité de vol, bien qu’il manque des données pour de nombreux groupes. La compression des structures trachéales est répandue, se produisant chez de nombreux insectes examinés. Cet arbre phylogénétique des Hexapodes a été modifié selon Misof et al. (2014) et Tihelka et al. (2021) (Modifié par B. GILLES) (Source : Harrison et al., 2023)
Développement embryologique des sacs aériens
Le développement des sacs aériens est étroitement lié au système trachéal, un réseau de tubes ramifiés qui transporte l’air directement aux tissus pénétrant par des spiracles situés sur l’exosquelette. Le processus de formation des sacs aériens commence au stade larvaire chez des insectes comme Drosophila. Les trachéoblastes, qui sont des cellules progénitrices spécifiques, migrent vers les trachées primaires au cours du développement embryonnaire, puis prolifèrent et se différencient en cellules épithéliales qui formeront les sacs aériens (Cabernard et Affolter, 2005).
Les mécanismes moléculaires sous-jacents à cette formation sont complexes. Par exemple, des récepteurs du facteur de croissance fibroblastique (FGF) régulent la migration des trachéoblastes et la formation des sacs. Ces récepteurs sont activés en réponse à des gradients de FGF exprimés dans les tissus environnants, dirigeant les cellules vers des régions où des structures respiratoires comme les sacs aériens sont nécessaires (Sato et Kornberg, 2002). En parallèle, le facteur de croissance épidermique (EGF) joue un rôle clé dans la régulation de la prolifération cellulaire, contrôlant l’augmentation du nombre de cellules qui formeront les sacs aériens et leur expansion (Cabernard & Affolter, 2005) (figure 2).
Figure 2 : A) Illustration schématique du développement des sacs aériens chez Drosophila – adaptée de (Sato & Kornberg, 2002) – B) Deux processus distincts de développement des sacs aériens (Source : Cabernard & Affolter, 2005)
Classification morphologique et fonctionnelle des sacs aériens
Les sacs aériens varient considérablement en taille et en structure selon les espèces, et ils peuvent être classés en trois types principaux, selon la présence ou l’absence de taenidies, qui sont des anneaux de renforcement des trachées :
Sacs taenidiaux : Les taenidies, sont présentes mais plus espacées, permettant une certaine flexibilité. Cette structure est typique chez les insectes ayant besoin d’une certaine rigidité trachéale tout en facilitant la compression et l’expansion (abeilles mellifères – Apis mellifera, criquet migrateur – Locusta migratoria, Odonates par exemple)
Sacs réticulés : Ils présentent des taenidies irrégulières formant un réseau en croisillons leur conférant une forme irrégulière. Ces sacs ont une capacité de compression accrue, idéale pour les insectes nécessitant des changements rapides de pression interne, comme ceux impliqués dans des cycles de respiration discontinue (Mouche domestique – Musca domestica, Guêpe sociale – Vespa vulgaris, ou le bourdon terrestre – Bombus terrestris par exemple)
Sacs ponctués : Ces sacs sont les plus compressibles et manquent complètement de taenidies. Ils permettent une grande variabilité de volume et sont souvent trouvés chez les insectes aquatiques ou ceux ayant des régimes respiratoires fluctuants (RRF)
Les RRF se réfèrent aux variations dans les modes de respiration alternant entre des phases actives, de repos ou d’échanges limités. Ces fluctuations s’observent lorsque les besoins en oxygène changent en fonction des activités métaboliques, de l’environnement ou de l’état physiologique de l’insecte.
Faucheux et Sellier (1971) ont proposé cette classification après avoir étudié la structure microscopique des sacs aériens chez plusieurs ordres d’insectes. Ces auteurs ont montré que la morphologie des sacs aériens est fortement influencée par les conditions écologiques dans lesquelles vit l’insecte.
Les régimes respiratoires fluctuants (RRF)
Un exemple de régime respiratoire fluctuant est le schéma de respiration discontinue (Discontinuous Gas Exchange, ou DGE), fréquemment observé chez les insectes terrestres comme les criquets et les coléoptères. Ce type de respiration comporte trois phases distinctes :
Phase fermée : Les spiracles (ouvertures du système respiratoire) restent fermés, empêchant tout échange de gaz avec l’environnement, ce qui permet de minimiser la perte d’eau.
Phase de flutter : Les spiracles s’ouvrent brièvement et partiellement, permettant à de petites quantités d’oxygène d’entrer sans une perte significative de dioxyde de carbone ou d’eau.
Phase ouverte : Les spiracles sont entièrement ouverts, permettant des échanges gazeux massifs, principalement pour expulser le dioxyde de carbone accumulé.
Ce type de régime respiratoire est particulièrement adapté aux environnements où les ressources sont limitées (par exemple, dans des conditions de sécheresse, où la conservation de l’eau est primordiale) ou lorsque l’insecte alterne entre des périodes de repos et d’activité intense, comme le vol ou la prédation.
Les sacs aériens, en particulier les sacs réticulés et ponctués, facilitent ces régimes respiratoires fluctuants en permettant des variations de volume qui régulent la pression interne et optimisent les échanges gazeux pendant ces différentes phases. Ces fluctuations de la respiration sont cruciales pour la gestion de l’oxygène et la conservation de l’eau, deux aspects essentiels de la survie dans des environnements difficiles ou lors d’activités métaboliquement exigeantes, comme le vol prolongé ou la recherche de nourriture.
Faucheux et Sellier (1971) ont proposé cette classification après avoir étudié la structure microscopique des sacs aériens chez plusieurs ordres d’insectes. Ces auteurs ont montré que la morphologie des sacs aériens est fortement influencée par les conditions écologiques dans lesquelles vit l’insecte.
Rôle des sacs aériens dans l’évolution du vol
Les insectes volants ont besoin de maintenir des apports d’oxygène élevés pour alimenter leurs muscles de vol, notamment les muscles thoraciques qui consomment une grande quantité d’énergie lors de la contraction rapide et soutenue nécessaire au vol (figure 3). Les sacs aériens ont probablement évolué en réponse à ces pressions sélectives. Leur fonction principale dans le cadre du vol est de permettre une ventilation par advection (transport d’une quantité d’un élément donné tel que lachaleur, l’énergie interne, unélément chimique par le mouvement (et donc la vitesse) du milieu environnant), augmentant ainsi le flux d’air à travers le système trachéal pour garantir un approvisionnement constant en oxygène (Harrison et al., 2019). Ils permettent également de réduire la densité corporelle de l’insecte, facilitant la manœuvrabilité et diminuant la charge alaire.
Figure 3 : Hypothèses sur la façon dont les sacs aériens affectent le fonctionnement du système trachéal – (A) Avec un hémocèle à compartiment unique et un système trachéal avec de grands sacs aériens plus conformes que les trachées, les changements de volume corporel entraînent l’advection à travers les sacs aériens et les trachées primaires, mais pas à travers les trachées distales – (B) Dans les systèmes trachéaux sans sacs aériens, les changements de volume corporel entraînent l’advection dans tout le système trachéal – (C) Les sous-régions créées par la valvulation fonctionnelle (créant plusieurs compartiments) peuvent permettre la compression des sacs aériens hautement conformes et des trachées moins conformes en réponse aux changements de volume corporel (Source : Harrison et al., 2023)
Chez des insectes comme les coléoptères, une forte variabilité est observée dans la présence de sacs aériens au sein même des familles. Par exemple, les coléoptères Scarabaeidae et Buprestidae, qui sont de puissants volants, possèdent des sacs aériens volumineux, tandis que les Carabidae, souvent non volants ou ayant une faible capacité de vol, en sont généralement dépourvus (Dittrich et Wipfler, 2021). Cependant, même parmi les Carabidae, certaines espèces volantes comme les cicendèles ont développé des sacs aériens thoraciques pour soutenir leur capacité de vol, soulignant la forte corrélation entre les sacs aériens et le vol.
Un autre exemple marquant est observé chez les Hyménoptères, en particulier les abeilles et les guêpes. Ces insectes possèdent des sacs aériens thoraciques et abdominaux bien développés qui leur permettent de voler sur de longues distances tout en maintenant une forte activité métabolique. Les abeilles, par exemple, utilisent leurs sacs aériens pour réduire la charge alaire et maximiser l’efficacité du vol lorsqu’elles collectent du nectar et du pollen (Taylor et al., 2010).
Exemples spécifiques
Chez le scarabée Dicronorrhina derbyana, des images de tomographie micro-CT montrent des sacs aériens massifs entourant les muscles de vol, illustrant leur rôle dans la réduction de la densité corporelle et dans l’amélioration de la maniabilité en vol (Harrison et al., 2019) (figure 4)
Les orthoptères tels que les criquets ont également des sacs aériens développés au niveau thoracique pour offrir de grandes capacités de vol lors de migrations sur de longues distances (Locusta migratoria). Le volume de ces sacs aériens augmente avec l’âge et la taille corporelle, avec une augmentation significative chez les adultes par rapport aux larves (Greenlee et al., 2009)
Figure 4 : Images de sacs aériens – (A) Tomodensitométrie (microCT) d’un scarabée (Dicronorrhina derbyana) – (B) Thorax disséqué d’un criquet (Locusta migratoria) – (C) Tête d’une fourmi mâle (Veromessor pergandei) – AS, sacs aériens ; FM, muscle de vol ; T, trachée ; FB, corps gras (Source : Harrison et al., 2023)
Sacs aériens et stratégies de vol
En plus de leur fonction de soutien à la respiration, les sacs aériens jouent également un rôle important dans la thermorégulation des insectes volants. Chez certains coléoptères, les sacs aériens thoraciques agissent comme des isolants thermiques, aidant à maintenir une température corporelle interne optimale pour le vol endothermique (producteur de chaleur) (Church, 1960). Cela est particulièrement utile pour les insectes volants dans des environnements froids ou lors de vols prolongés, durant lesquels l’évitement de la surchauffe des muscles est crucial.
Les sacs aériens sont également impliqués dans la réduction du coût énergétique du vol. En remplaçant le tissu dense par des cavités remplies d’air, les sacs aériens allègent la structure corporelle de l’insecte, ce qui réduit les besoins énergétiques pour maintenir le vol stationnaire ou prolongé (Newport, 1851). Cependant, il est important de noter que cette réduction de densité peut être contre-productive dans certains cas. Par exemple, une augmentation du volume corporel sans augmentation de masse peut augmenter la traînée aérodynamique lors du vol à grande vitesse (résistance au vent sans gain de puissance musculaire). C’est pourquoi les sacs aériens doivent être soigneusement régulés pour maximiser l’efficacité du vol sans entraîner de pénalités de performance : compression ou expansion (musculaire ou passive) des sacs aériens en fonction des activités et des besoins.
Par exemple, lors d’un vol soutenu chez les bourdons et les abeilles, les sacs aériens thoraciques se dilatent lorsque l’insecte est en vol stationnaire pour maximiser l’oxygénation des muscles de vol, mais ils se compressent partiellement lors de vols rapides pour réduire la traînée (Harrison et al., 2019) (vidéo ci-dessous).
Diversité des sacs aériens et sélection naturelle
La distribution phylogénique des sacs aériens montre clairement qu’ils ont évolué plusieurs fois de manière indépendante chez divers ordres d’insectes.
Cette convergence évolutive illustre l’importance sélective de ces organes pour le vol et d’autres adaptations comme la régulation de la flottabilité.
Les sacs aériens ne sont pas seulement bénéfiques pour les insectes volants, mais ont également permis des innovations évolutives dans la taille corporelle. Par exemple, les coléoptères à « cornes » comme le Rhinocéros (Oryctes nasicornis) ou le Lucane cerf-volant (Lucanus cervus)) ou encore les mouches Diopsidae (vidéo ci-dessous) dont les yeux sont perchés à l’extrémité de longs pédoncules à la manière des requins-marteaux, possèdent des sacs aériens dans leurs cornes, ce qui réduit leur poids sans compromettre leur taille, permettant ainsi la sélection de ces structures sexuelles hypertrophiées (McCullough et Tobalske, 2013).
Extension des pédoncules d’une Diopsidae à l’aide des sacs aériens au niveau de la tête
L’évolution des sacs aériens montre donc un lien fort entre l’optimisation du vol, la réduction de la densité corporelle et les contraintes environnementales. Les insectes ayant besoin de vols prolongés, comme les pollinisateurs ou les insectes migrateurs, ont évolué pour maximiser l’utilisation de ces structures respiratoires complexes, tandis que ceux ayant des besoins respiratoires moindres, comme les insectes terrestres non volants, les ont progressivement perdus.
Compression trachéale : un mécanisme complémentaire
Dans les cas où les sacs aériens sont absents ou sous-développés, les insectes peuvent compenser en utilisant la compression trachéale pour faciliter la ventilation. Ce processus implique une compression des trachées secondaires et tertiaires, souvent à l’aide de mouvements abdominaux. Ce mécanisme permet d’entretenir un flux d’air constant dans le système trachéal sans avoir besoin de structures comme les sacs aériens (Socha et DeCarlo, 2008).
Un exemple notable de ce phénomène s’observe chez les coléoptères carabidés, qui sont généralement des marcheurs terrestres. Ces insectes n’ont pas de sacs aériens développés, mais ils compressent activement leurs trachées pour augmenter le flux d’air pendant l’activité locomotrice intense (Socha et al., 2008) (figure 5). Leurs trachées sont souvent ovales en coupe transversale, permettant une flexibilité accrue lors de la compression, un élément clé pour leur survie dans des environnements nécessitant une mobilité rapide.
Figure 5 : Dispositif expérimental utilisé pour visualiser la compression trachéale interne du carabe Pterostichus stygicus. Les images radiographiques montrent ici le tube trachéal (flèche bleu) contracté et dilaté dans le mésothorax entre le premier et le deuxième coxae (articulation entre la patte et le thorax (Modifié par B. GILLES) (Source : Socha et al., 2008)
Avantages et inconvénients des sacs aériens
Les avantages des sacs aériens sont nombreux, notamment pour les insectes volants. Cependant, ces avantages ont des contreparties importantes. L’un des principaux coûts évolutifs de leur présence est la réduction de la capacité à stocker des réserves d’eau et de nutriments. Ceci est particulièrement crucial dans les environnements secs où les insectes doivent optimiser la rétention d’eau pour survivre (Bradley et al., 1999).
Dans les environnements arides, les insectes ayant de grands sacs aériens peuvent être désavantagés car ils manquent de réserves d’eau suffisantes pour compenser les pertes dues à l’évaporation. En revanche, les insectes plus petits et ceux ayant des structures trachéales plus rigides (moins de sacs aériens) sont mieux équipés pour limiter ces pertes.
Innovations évolutives permises par les sacs aériens
Les tibias d’orthoptères tels que ceux des criquets renferment également des sacs aériens, réduisant la masse des pattes et facilitant ainsi la locomotion (figure 6). Cette adaptation montre que le rôle des sacs aériens n’est pas limité aux fonctions respiratoires, mais qu’ils participent aussi à l’allègement du squelette et à l’optimisation de la locomotion (Ruan et al., 2018).
Figure 6 : Réseau trachéal dans une patte postérieure de criquet (Tridactylidae) et sacs aériens (As) dans le tibias (photo à gauche et croquis à droite) (Source : Ruan et al., 2018)
Adaptations au milieu aquatique et fonction des sacs aériens
Chez les insectes aquatiques, les sacs aériens sont relativement rares ou limités. En raison de la pression hydrostatique élevée sous l’eau, leur présence peut rendre difficile le maintien de la position sous l’eau. La flottabilité excessive est un problème majeur pour les insectes aquatiques. C’est pourquoi les larves de nombreuses espèces, comme celles des libellules, ont des systèmes trachéaux fermés sans sacs aériens (Tillyard, 1917) (figure 7).
Cependant, certains insectes aquatiques, comme les larves de Chaoborus, ont développé des sacs aériens spécialisés qui leur permettent de réguler leur flottabilité. Ces sacs aériens plus résistants à la pression permettent aux larves de contrôler leur position dans la colonne d’eau, une capacité essentielle pour chasser les zooplanctons et échapper aux prédateurs (McKenzie et al., 2022) (figure 7 et 8).
Figure 7 : Présence et fonction des sacs aériens dans les systèmes trachéaux des larves d’insectes aquatiques. Le bleu clair représente les sacs aériens, le bleu foncé représente les trachées principales. Les volumes variables des sacs aériens chez Mochlonyx et Chaoborus leur permettent d’atteindre une flottabilité neutre pour flotter au milieu de l’eau. En revanche, les nymphes d’Eucorethra et de libellules n’ont pas de sacs aériens et ont une flottabilité négative. Eristalis arbustorum possède quant à lui des sacs aériens qui fonctionnent pour déplacer l’air par advection à travers leur long tube respiratoire postérieur. L’inhalation ou l’expiration d’air leur permet de varier leur flottabilité à volonté. Sacs aériens de Chaoborus basés sur Bardenfleth et Ege (1916) ; silhouette d‘Eristalis basée sur Dunavan (1929) (Modifié par B. GILLES) (Source : Harrison et al., 2023)Figure 8 : Les larves de moucherons aquatiques Chaoborus sont les seuls insectes capables de contrôler leur flottabilité. Leurs sacs aériens trachéaux agissent comme un moteur mécanochimique alimenté par le pH. Crédit : Joyce Gross-2018) (Source : Bugguide.net)
Régulation de la ventilation trachéale : l’importance des mouvements abdominaux et des cycles de compression
La ventilation trachéale chez les insectes repose sur des mécanismes actifs et passifs. Chez les insectes de grande taille ou à métabolisme élevé, tels que les criquets et les coléoptères, les mouvements rythmiques de l’abdomen ou du thorax provoquent la compression des sacs aériens et des trachées, ce qui génère un flux d’air constant (Westneat et al., 2003).
Les insectes possédant des sacs aériens volumineux, comme certains hyménoptères, montrent des cycles de compression-expansion qui permettent de réguler l’apport d’oxygène et l’élimination du dioxyde de carbone. Ce type de ventilation est particulièrement important lors des périodes de vol intense ou de stress thermique, lorsque la demande en oxygène est maximale.
Conclusion
Les sacs aériens sont des structures adaptatives cruciales qui ont permis aux insectes de développer une grande diversité de stratégies écologiques et physiologiques. De leur rôle dans la respiration et le vol à leurs contributions à la régulation de la flottabilité et à l’évolution de structures complexes (musculatures, appendices hypertrophiés, systèmes de contrôle de la flottabilité, etc.), les sacs aériens ont profondément influencé l’évolution de ces arthropodes.
Leur présence n’est pas seulement une réponse aux besoins respiratoires mais aussi une innovation permettant des stratégies complexes d’adaptation à l’environnement. Cependant, les sacs aériens posent des défis évolutifs, notamment en termes de gestion de l’eau (transpiration, déshydratation, etc.) et de la compression sous pression hydrostatique. Des recherches futures pourraient se concentrer sur l’exploration de la diversité fonctionnelle des sacs aériens en lien avec les stratégies de survie des insectes dans des environnements extrêmes.
Bibliographie
Bradley, T. J., Williams, A. E. & Rose, M. R. (1999) : Physiological responses to selection for desiccation resistance in Drosophila melanogaster. Am. Zool. 39, 337-345 (lien)
Cabernard C. & Affolter M. (2005) : Distinct roles for two receptor tyrosine kinases in epithelial branching morphogenesis in Drosophila. Dev. Cell. 9,831-842 (lien)
Church, N. S. (1960) : Heat loss and the body temperatures of flying insects : II. heat conduction within the body and its loss by radiation and convection. J. Exp. Biol. 37, 186-212 (lien)
Dittrich, K. & Wipfler, B. (2021) : A review of the hexapod tracheal system with a focus on the apterygote groups. Arthropod. Struct. Dev. 63, 101072 (lien)
Faucheux, M. J. & Sellier, R. (1971) : L’ultrastructure de l’intima cuticulaire des sacs aeriens chez les Insectes. C. R. Hebd. Séances Acad. Sci. D 272D, 2197-2200
Greenlee, K. J. et al. (2009) : Synchrotron imaging of the grasshopper tracheal system: morphological components of tracheal hypermetry and the effect of age and stage on abdominal air sac volumes and convection. Am. J. Physiol. Comp. Regul. Integr. Physiol. 297 (lien)
Guha A. & Kornberg T.B. (2005) : Tracheal branch repopulation precedes induction of the Drosophila dorsal air sac primordium. Dev. Biol. 287, 192-200 (lien)
Harrison, J. F. et al. (2019) : Physiological responses to gravity in an insect. Proc. Natl. Acad. Sci. USA 117, 2180-2186 (lien)
Herhold H.W. ; Davis S.R. ; DeGrey SP.P & Grimaldi D.A. (2023) : Comparative anatomy of the insect tracheal system part 1 : introduction, Apterygotes, Paleoptera, Polyneoptera. Bull. Am. Mus. Nat. Hist. 459, 1-184 (lien)
Lee M.O. (1929) : The function of the air sacs in holopneustic insects. Science 69, 334-335 (lien)
Mccullough, E. L. & Tobalske, B. W. (2013) : Elaborate horns in a giant rhinoceros beetle incur negligible aerodynamic costs. Proc. R. Soc. B 280, 2013019 (lien)
Mckenzie, E. K. G., Kwan, G. T., Tresguerres, M. & Matthews, P. G. D. (2022) : A pH-powered mechanochemical engine regulates the buoyancy of Chaoborus midge larvae. Curr. Biol. 32, 927-933.e5 (lien)
Newport, G. (1851) : On the formation and use of air sacs and dilated trachea in insects. J. Linn. Soc. 20, 419
Ruan, Y., Li, Y., Zhang, M., Chen, X., Liu, Z., Wang, S. & Jiang, S. (2018) : Visualisation of insect tracheal systems by lactic acid immersion. J. Microsc. 271, 230-236 (lien)
Sato, M. & Kornberg, T. B. (2002) : FGF is an essential mitogen and chemoattractant for the air sacs of the Drosophila tracheal system. Dev. Cell 3, 195-207 (lien)
Socha, J. J. & Decarlo, F. (2008) : Use of synchrotron tomography to image naturalistic anatomy in insects. SPIE 2008, 70780A (lien)
Swammerdam J.J. (1737) : Bybel der Nature. Leyden: Severin
Taylor, R. A. J., Bauer, L. S., Poland, T. M. & Windell, K. N. (2010) : Flight performance of Agrilus planipennis (Coleoptera: Buprestidae) on a flight mill and in free flight. J. Insect Behav. 23, 128-148 (lien)
Tillyard, R. J. (1917) : The Biology of Dragonflies (Odonata or Paraneuroptera). Cambridge: Cambridge University press
Westneat, M. W., Betz, O., Blob, R. W., Fezzaa, K., Cooper, W. J. & Lee, W. K. (2003) : Tracheal respiration in insects visualized with synchrotron X-ray imaging. Science 299, 558-560 (lien)
Tout au long de leur cycle de vie – allant de l’œuf jusqu’à à la fin de leur vie d’adulte sexué – les insectes font face à de nombreuses menaces, aussi bien d’origine abiotique (e. g. température, vent, etc.) que biotique (e.g. ennemis naturels). Ils ont développé un incroyable arsenal de défenses pour se protéger des pressions qu’ils subissent et qui les menacent. Parmi les différentes barrières de défenses, c’est le système immunitaire qui est le point focal de cet article.
Souvent décrit comme le dernier rempart pour se défendre, le système immunitaire des insectes a longtemps été considéré comme moins sophistiqué que celui des vertébrés. Cependant, depuis les années 1990, les études montrant sa complexité se sont multipliées. Jules Hoffmann (directeur de recherche émérite, CNRS, Strasbourg) a d’ailleurs obtenu – avec Bruce Beutler et Ralph Steinman – le prix Nobel de physiologie et de médecine, en 2011, pour son travail sur l’immunité innée de la Drosophile.
Description générale du système immunitaire
Le système immunitaire latosensu représente l’ensemble des processus biologiques qui protègent un organisme des maladies résultant des agents pathogènes et des cellules cancéreuses, en les identifiant et les éliminant. Sa fonction n’est donc pas seulement l’exclusion des parasites et la lutte contre les infections ! Il permet de distinguer un large éventail d’agents infectieux en les différenciant des cellules saines et des tissus hôtes.
Le système immunitaire des invertébrés répond, comme celui des vertébrés, de façon innée. En revanche, il se distingue de celui des vertébrés par son absence d’immunité adaptative avec une mémoire basée sur des immunoglobulines spécifiques (protéines douées d’activités d’anticorps) (Carton & Nappi 2001). Cependant, chez les insectes, le système immunitaire inné livre une réponse humorale (la prophénoloxidase et les peptides antimicrobiens, voir section réponse immunitaire) et une réponse cellulaire (hémocytes, voir sections suivantes) dont le déclenchement dépent de la menace détectée. De nombreux organes sont impliqués dans le déclenchement de cette réponse, comme par exemple le corps gras et l’intestin moyen (figure 1) (lien).
Figure 1 : Anatomie du système immunitaire. La cavité corporelle de l’insecte, appelée hémocoele, est un espace fluide et dynamique qui abrite des tissus ayant une activité immunitaire. Les hémocytes sont présents dans la circulation (hémocytes circulants) et attachés aux tissus (hémocytes sessiles). Le corps gras (fat body), l’intestin moyen (midgut), les glandes salivaires (salivary glands) et d’autres tissus produisent de nombreux facteurs immunitaires (d’après Hillyer, 2016)
Activation du système immunitaire
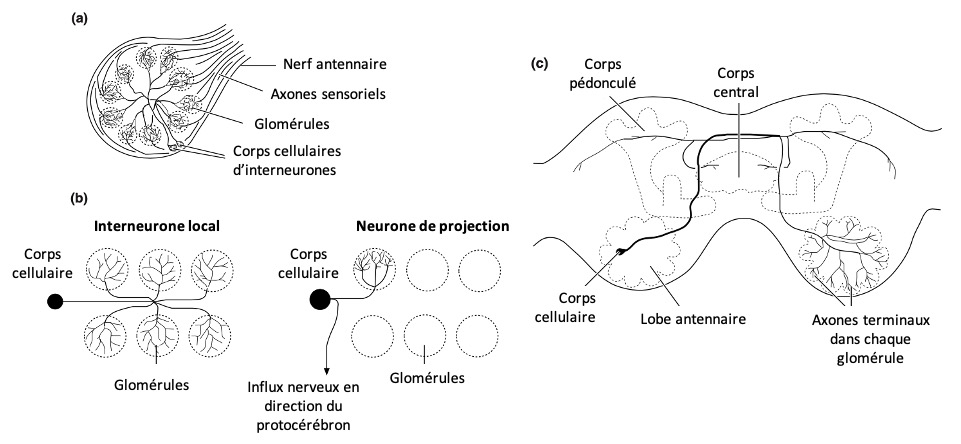
La réponse immunitaire des insectes est activée par la reconnaissance d’un large spectre de motifs du non-soi, et notamment, par ces cellules immunitaires appelées hémocytes (l’équivalent de nos globules blancs ou leucocytes). Ce non-soi est reconnu comme étranger grâce à des molécules spécifiques à la surface de ses cellules (appelés les modèles moléculaires associés aux agents pathogènes = Pathogen Associated Molecular Patterns ou PAMPs). Les molécules du pathogènes sont reconnues par leur équivalent présents à la surface des hémocytes, appelés peptides de reconnaissance des formes (Pattern Recognition Peptides = PRPs) (figure 2) (Siva-Jothy et al. 2005).
Une fois le pathogène détecté, une cascade de réponses est déclenchée en fonction de la menace (ex : bactérienne, virale, etc.), adaptant ainsi la réponse immune.
Exemple de l’encapsulation* d’un œuf de parasitoïde
Dans le cas d’une infection suite à l’injection d’un œuf de parasitoïde dans l’organisme hôte (œuf ou chenille par exemple), les hémocytes circulant dans l’hémolymphe vont immédiatement sonner l’alarme (Siva-Jothy et al. 2005). Cela entraine le recrutement d’autres hémocytes qui vont s’agrègent autour de l’œuf afin de l’isoler du reste du corps de l’insecte (figures 2, 3). Le recrutement des hémocytes est rapide et peut durer dans le temps.
Par exemple, chez la Cochylis de la vigne (Eupoecilia ambiguella), l’augmentation des hémocytes est significative dès 2h après une injection mimant une attaque de parasitoïde, et augmente jusqu’à 24h post-infection (Vogelweith, données non publiées). Les hémocytes vont en même temps et tout aussi rapidement entrainer l’activation de la cascade enzymatique de la prophénoloxydase (Cerenius & Soderhall 2004). L’activation de cette enzyme va entrainer la production de mélanine et former une capsule mélanique autour de l’œuf (figures 2, 3). L’activation de la cascade entraine également la production de molécules cytotoxiques et de quinones qui vont être toxiques pour l’œuf et participer à son élimination (Nappi & Ottaviani 2000). Il faut noter que l’enzyme prophénoloxidase est également impliquée dans d’autres processus tels que la coagulation et la sclérotinisation (processus aboutissant à une cuticule rigide chez les arthropodes) après une blessure, permettant de la refermer (Sugumaran et al. 2000).
*Encapsulation : mécanisme de défense en réponse à l’invasion d’un organisme étranger à l’intérieur d’un insecte. Ainsi, l’hôte forme une capsule autour de l’organisme étranger grâce aux hémocytes et à la production de mélanine, et l’isole du reste du corps.
Suite à une infection microbienne, un ensemble de peptides antimicrobiens (AMPs) est produit et sécrété dans l’hémolymphe par les hémocytes (figure 2), le corps gras et les épithéliums (Eleftherianos et al. 2021). A l’heure actuelle, plus de 360 AMPs ont été décrits et isolés chez les insectes avec des propriétés variés aussi bien antibactériennes, antifongiques qu’antiparasitaires (voir la revue de Eleftherianos et al. 2021). Contrairement aux autres effecteurs (hémocytes et prophénoloxidase), cette ligne de défense est plus spécifique et très efficace pour neutraliser les microorganismes (e.g. bactéries). Cependant, sa mise en œuvre est relativement lente en raison de son caractère inductible de cette voie immunitaire (Haine et al. 2008). Une fois stimulée, la production de peptides antimicrobiens peut être importante et se prolonger relativement longtemps compte tenu de la durée de vie des invertébrés (Haine et al. 2008).
La réponse immunitaire des insectes a donc une action humorale et cellulaire, immédiate grâce aux hémocytes et à la Phénoloxydase (PO – mélanisation des agents infectieux) (appelée immunité constitutive), et à long terme par l’intermédiaire des peptides antimicrobiens.
Le système immunitaire et ses compromis
Malgré cet outil performant, les insectes ne sont évidemment pas pour autant totalement protégés. En effet, maintenir une immunité constitutive et mettre en place une réponse immunitaire sont des fonctions biologiques couteuses. L’insecte n’ayant pas accès à des ressources illimitées, des compromis sont donc indispensables que ce soit au sein du système immunitaire lui-même ou par le biais d’autres traits d’histoire de vie, tels que la croissance ou la reproduction.
En effet, un compromis entre l’immunité constitutive (hémocytes et PO) et l’immunité à plus long terme (peptides antimicrobiens), modulé par la nourriture, a été mis en évidence chez les chenilles de la Cochylis de la vigne (E. ambiguella) (figure 4a) (Vogelweith et al. 2011) et de la Noctuelle méditerranéenne (Spodoptera littoralis) (Cotter et al. 2011). Ainsi, les chenilles de la Cochylis (figure 4b) se nourrissant du cépage savagnin rose (Gewürztraminer) seraient plus à même de se défendre contre les parasitoïdes (plus d’hémocytes et de prophénoloxidase) que celles élevées sur un chasselas qui elles seraient plus armées contre les bactéries (plus d’activité antimicrobienne) (figure 4c) (Vogelweith et al. 2011).
Un autre exemple de compromis, cette fois entre différents traits d’histoire de vie, est celui observé chez le Puceron vert du pois (Acyrthosiphon pisum), où les individus les plus féconds sont les moins résistants à une attaque par un parasitoïde (Gwynn et al. 2005). Ces insectes doivent donc adapter leur « investissement » dans l’immunité en fonction de leur environnement, de leur mode de vie (e.g. grégaire, solitaire) et des pressions qu’ils subissent.
C’est en raison de la complexité de ces interactions qu’est née une discipline appelée l’« immuno-écologie ».
Les hémocytes sont des cellules très nombreuses, de morphologie variable, circulant librement dans l’hémolymphe des insectes (figures 1, 2, 3). Ils proviennent des tissus hématopoïétiques (e.g. corps gras, intestin moyen, glandes salivaires) dans lesquels ils se différencient et se divisent. Ils présentent différents types spécialisés dans des fonctions particulières de l’immunité (e.g. reconnaissance du non-soi), et dont le nombre varie en fonction du stade de développement de l’individu et de l’espèce (Lavine & Strand 2002).
Par exemple, chez l’Eudémis de la vigne (Lobesia botrana), 5 types d’hémocytes sont présents chez les jeunes chenilles puis 4 types chez les chenilles au dernier stade (figure 5). Certains hémocytes comme les plasmatocytes ou les granulocytes sont responsables de la reconnaissance des corps étrangers (figure 5) : les plasmatocytes sont les premiers à adhérer à un œuf de parasitoïde par exemple. Outre leur rôle dans l’encapsulation (figures 2, 3), les hémocytes sont également impliquées dans la phagocytose (processus consistant à l’internalisation par les hémocytes de petites particules étrangères conduisant à leur dégradation) des corps étrangers, la nodulation des bactéries (micro-agrégation d’hémocytes autour de plusieurs microorganismes infectieux, bactéries, etc.) et la coagulation de l’hémolymphe suite à une blessure dans la cuticule (Lavine & Strand 2002; Siva-Jothy et al. 2005). Ils constituent donc un paramètre clé de la réponse immunitaire (voir Lavine & Strand 2002 pour une description approfondie).
Figure 5 : Photographies et schémas annotés des différents types d’hémocytes d’une chenille L. botrana, au dernier stade larvaire : granulocytes (GR ; a, e), plasmatocytes (PL ; b, f), oenocytoïdes (OE ; c, g), spherulocytes (SP ; d, h). Sur les schémas annotés, « C » indique le cytoplasme, « G » les granules, « M » la membrane, « N » le noyau et « S » les sphérules (d’après Vogelweith et al. 2016)
Bibliographie
Carton, Y., Poirié, M., & Nappi, A. J. (2008) : Insect immune resistance to parasitoids. Insect Science, 15(1), 67–87 (lien)
Cerenius, L., & Söderhäll, K. (2004) : The prophenoloxidase-activating system in invertebrates. Immunological Reviews, 198, 116–126 (lien)
Cotter, S. C., Simpson, S. J., Raubenheimer, D., & Wilson, K. (2011) : Macronutrient balance mediates trade-offs between immune function and life history traits. Functional Ecology, 25(1), 186–198 (lien)
Eleftherianos, et al. (2021) : Diversity of insect antimicrobial peptides and proteins – A functional perspective: A review. In International Journal of Biological Macromolecules (Vol. 191, pp. 277–287). Elsevier B.V. (lien)
Gwynn, D. M., Callaghan, a, Gorham, J., Walters, K. F. a, & Fellowes, M. D. E. (2005) : Resistance is costly: trade-offs between immunity, fecundity and survival in the pea aphid. Proceedings. Biological Sciences / The Royal Society, 272(1574), 1803–1808 (lien)
Haine, E. R., Moret, Y., Siva-Jothy, M. T., & Rolff, J. (2008) : Antimicrobial defense and persistent infection in insects. Science, 322, 1257–1259 (lien)
Hillyer, J. F. (2016). Insect immunology and hematopoiesis. Developmental and Comparative Immunology (lien)
Lavine, M. D. D., & Strand, M. R. R. (2002) : Insect hemocytes and their role in immunity. Insect Biochemistry and Molecular Biology, 32(10), 1295–1309 (lien)
Nappi, A. J., & Carton, Y. (2001) : Immunogenetic aspects of the cellular immune response of Drosophila against parasitoids. Immunogenetics, 52(3–4), 157–164 (lien)
Nappi, A. J., & Ottaviani, E. (2000) : Cytotoxicity and cytotoxie molecules in invertebrates. BioEssays, 22, 469–480 (lien)
Siva-Jothy, M. T., Moret, Y., & Rolff, J. (2005) : Insect Immunity : An Evolutionary Ecology Perspective. Advances In Insect Physiology, 32(05), 1–48 (lien)
Strand, M. R. (2008) : The insect cellular immune response. Insect Science, 15(1), 1–14 (lien)
Sugumaran, M., Nellaiappan, K., & Valivittan, K. (2000) : A new mechanism for the control of phenoloxidase activity: inhibition and complex formation with quinone isomerase. Archives of Biochemistry and Biophysics, 379(2), 252–260 (lien)
Vogelweith, F. (2013) : Effet de la plante hôte sur l’immunité des insectes phytophages : Influence du cépage de vigne sur la capacité des vers de la grappe à se défendre contre des bio-agresseurs. University of Burgondy (lien)
Vogelweith, F., Moret, Y., Monceau, K., Thiéry, D., & Moreau, J. (2016) : The relative abundance of hemocyte types in a polyphagous moth larva depends on diet. Journal of Insect Physiology, 88, 33–39 (lien)
Vogelweith, F., Thiéry, D., Quaglietti, B., Moret, Y., & Moreau, J. (2011) : Host plant variation plastically impacts different traits of the immune system of a phytophagous insect. Functional Ecology, 25(6), 1241–1247 (lien)
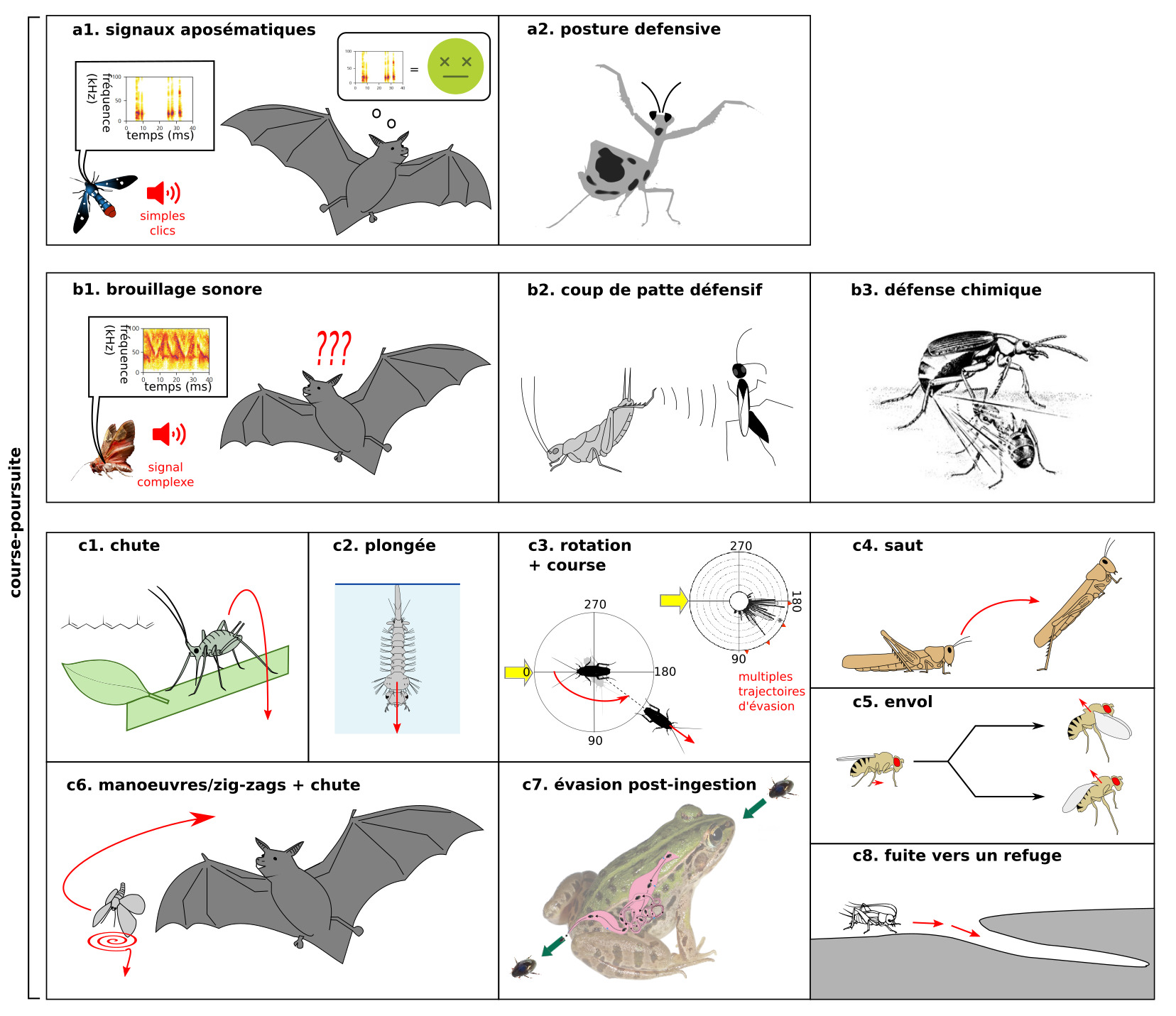
Cet article sur la neuroécologie des courses-poursuites est divisé en deux parties. Ne manquez pas la première partie sur l’écologie des jeux de cache-cache/évasion, avant de lire celle-ci, qui porte plus sur des aspects neuroéthologiques.
II – Détection et traitement des informations sensorielles
Les prédateurs produisent des indices (cues) qui peuvent être détectés par la proie. La réponse d’évasion est décomposable en trois parties, comme décrit dans (1). Il s’agit tout d’abord d’une classification de l’information sensorielle incluant des composants perceptuels (nature du stimulus), et de valeur (est-ce que le stimulus est menaçant ou non). Ensuite, il y a une phase de sélection d’action (initiation de la réponse), autrement dit quelle action du répertoire comportemental est la plus appropriée à la situation. Enfin, il y a une phase d’action (exécution de la réponse).
Après une description du « cahier des charges » d’un système de fuite, nous décrirons certains circuits neurosensoriels connus dans la détection des prédateurs.
A) Caractéristiques générales des réponses d’évasion
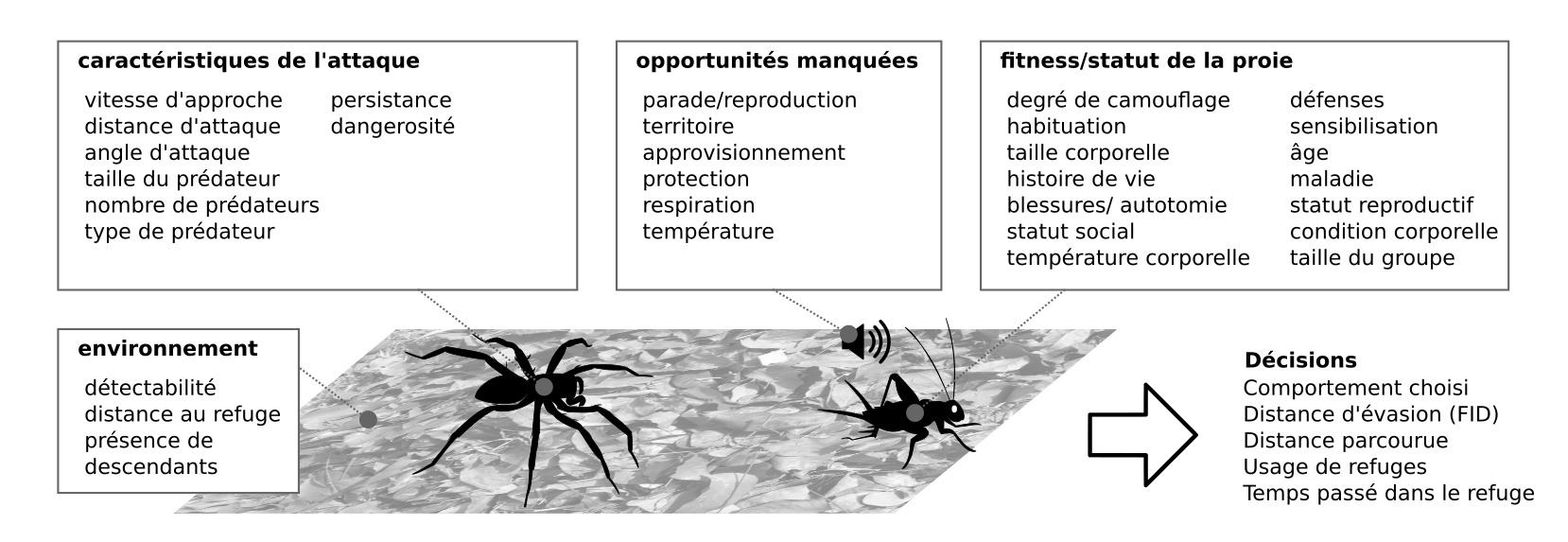
Une interaction physique avec un prédateur conduit à une perte de fitness. Dans le cas le plus extrême, l’interaction proie-prédateur conduit à la mort de la proie. Même si la proie parvient à s’échapper, les blessures se traduisent par exemple par un perte de la capacité locomotrice pour rechercher un partenaire ou de la nourriture, ou par un handicap lors de combat avec des conspécifiques (1). Les pressions de sélections sur l’évasion se traduisent par des systèmes nerveux innés (hardwired) avec de hauts niveaux d’efficacité, avec toutefois une part de flexibilité.
Les circuits d’évasion (2) :
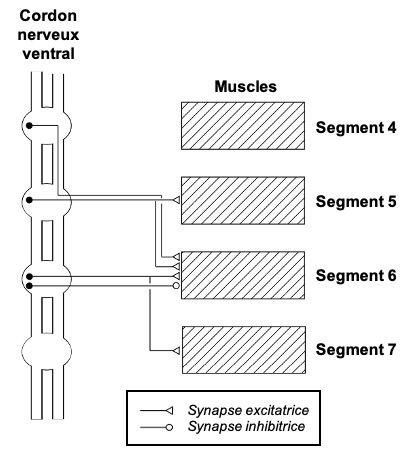
Sont rapides et avec un timing précis. Une milliseconde peut faire la différence. Chez les insectes, il n’existe pas de gaine de myéline (pas de conduction saltatoire). La vitesse de conduction dépend alors du diamètre des neurones : plus le neurone est gros, plus rapide est la conduction de l’influx nerveux. On retrouve des neurones géants impliqués dans l’évasion chez de nombreux ordres d’insectes
Sont résistants aux faux négatifs et aux faux positifs, et intégrés à la biologie. S’échapper dans des situations inappropriées peut être très coûteux (figure 4)
Sont matched. Les circuits sont adaptés aux forces et faiblesses du prédateurs. Les structures neurales sont coûteuses à entretenir, et posséder des systèmes sensoriels périphériques sélectifs aux stimuli les plus pertinents pour la biologie permet de s’affranchir de calculs neuronaux supplémentaires (3). Par exemple, les papillons de nuit présentent des tympans sélectifs à une gamme de hautes fréquences qui correspondent grosso modo à celle utilisées par les chauves-souris (4). L’entretien des systèmes nerveux est coûteux, et on a observé une sensibilité plus forte des neurones AN2 (des neurones clés dans la perception des ultrasons) chez les populations de grillons exposés à un fort risque de prédation par des chauve-souris, en comparaison avec une autre population vivant dans une zone avec un risque plus bas (5)
Présentent souvent une parcimonie neuronale, la rapidité de la réponse impose une limite numérique sur le nombre de neurones
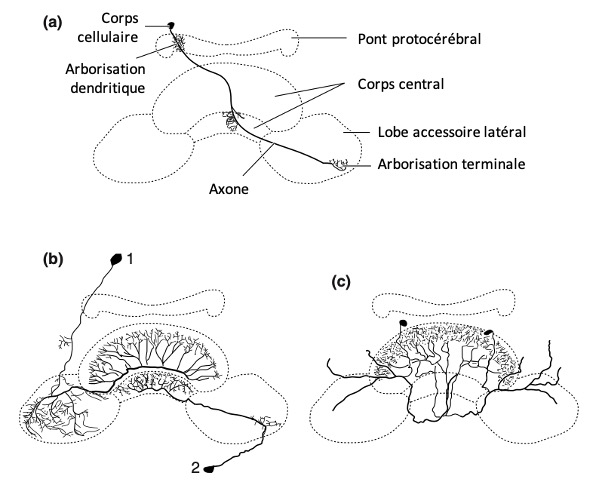
Sont innés et adaptatifs. Se baser entièrement sur l’expérience du danger est risqué. Certains motifs sont extrêmement conservés, comme par exemple les neurones sensibles à un stimulus visuel looming (voir figure 5.1 ci-dessous). On décrit souvent l’évasion comme de simples boucles réflexes, mais il s’agit en fait d’une réponse coordonnée décomposable en une série de « sous-comportements » (6)
On parle de fixed action pattern pour décrire un comportement stéréotypé, caractéristique d’une espèce et déclenché par un stimulus releaser.Cependant, certaines réponses d’évasion peuvent être interrompus en cours de route, et peuvent dépendre de l’expérience, ce qui n’est pas le cas d’un fixed action pattern au sens strict.
Figure 4 : Aspects multifactoriels de l’évasion : nombreux facteurs internes et externes influencent les décisions prises lors de l’évasion (Source : P. Clémençon)
B) Système visuel de détection des collisions
Lorsqu’un prédateur s’approche d’une proie, cela produit une tâche (projection rétinienne) dont la taille angulaire ϴ(t) augmente de manière non-linéaire au cours du temps (figure 5c). Celle-ci ne dépend que du ratio l/v entre la demi-taille de l’objet et sa vitesse d’approche. On appelle cela le looming (figure 5.1). De nombreux neurones sensibles au looming ont été découvert chez les Vertébrés et Invertébrés, soulignant une convergence évolutive et l’importance de ces indices pour la biologie des animaux (10) (lien).
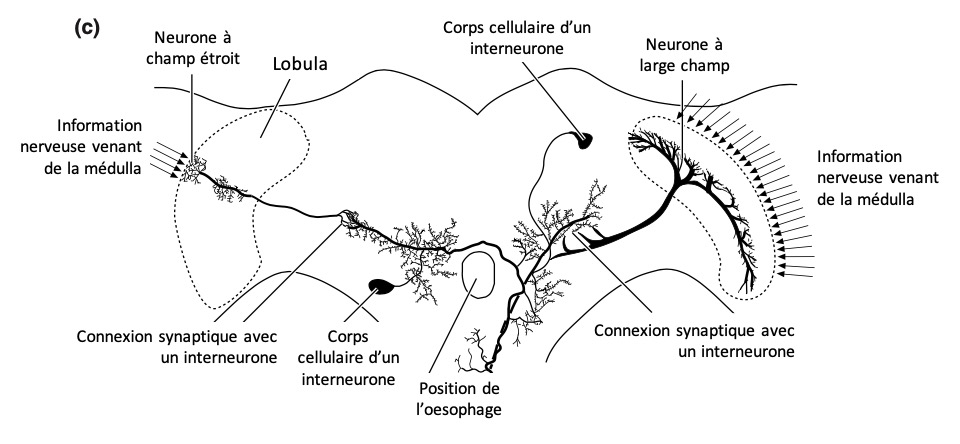
Le système visuel des insectes est assez bien connu (lien). Les lobes optiques sont divisés en plusieurs régions. Il y a dans l’ordre : les photorécepteurs, la lamina, la médulla puis la lobula/lobula plate. Le traitement de l’information visuelle se fait de manière progressive (figure 6). Les cellules plus périphériques encodent la luminance d’un objet, celles des couches inférieures encodent les variations de luminance, et les cellules de couches encore inférieures encodent des caractéristiques visuelles plus fines, comme le mouvement (Elemenary Motion Detectors EMD).
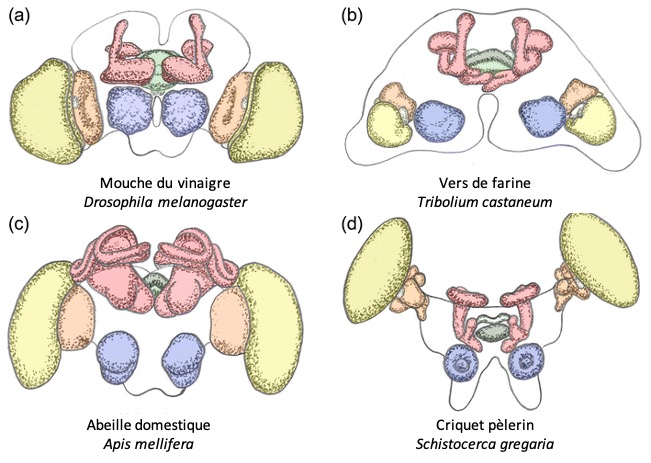
En raison de la présence d’outils génétiques, et de son statut d’« organisme modèle », l’anatomie du système visuel de détection des collisions est plus connu chez la Drosophile que chez le criquet.
Chez la Drosophile, on observe deux types de réponses. Une réponse courte d’urgence, qui est rapide (chaque ms compte !) mais qui nécessitera des manœuvres pour corriger la trajectoire du vol instable, et une réponse longue, durant laquelle on observe une élévation des ailes avant la saut (préparation de la posture) (7).
Les neurones clés sont les cellules LPLC2, LC4, et les neurones descendants (DN), le plus célèbre étant la Fibre Géante (GF=Giant Fiber). Les cellules LPLC2 reçoivent des entrées des neurones T4 et T5 qui sont sensibles au mouvement. Les cellules LPLC2 indiquent la taille de l’objet, alors que les cellules LC4 (Lobula Columnar type 4) indiquent la vitesse de l’objet. On parle de multiplexing d’information (ou de traitement parallèle) : les caractéristiques d’un prédateur (sa taille et sa vitesse d’approche) sont traitées en même temps par des circuits spécialisés parallèles, accélérant le traitement.
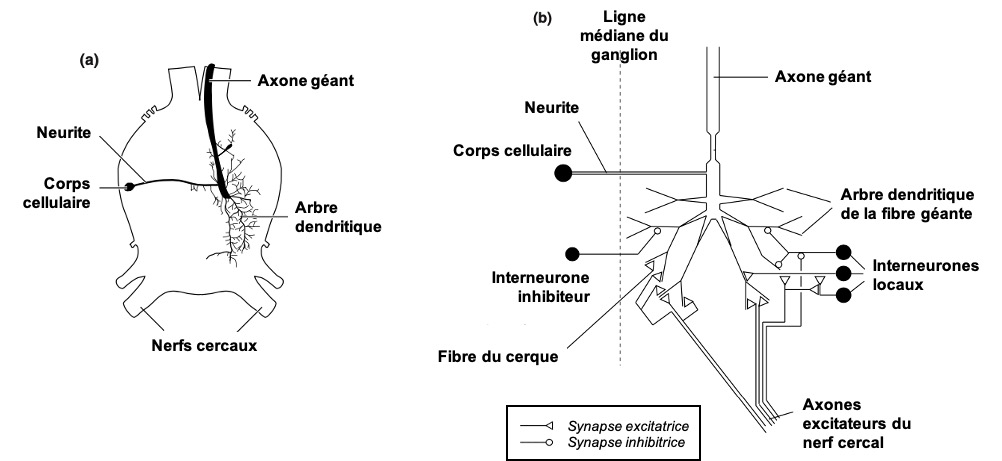
La Fibre Géante réaliserait l’opération de multiplication l/v en combinant les entrées synaptiques provenant des cellules LPLC2 (taille, cellule orange sur la figure 6), et celles provenant des cellules LC4 (vitesse, cellule bleue sur la figure 6) (11). Lorsqu’un potentiel d’action est produit par la Fibre Géante, il se produit toujours des évasions de type « mode-court », sans élévation des ailes au préalable. La Fibre Géante est souvent décrite comme un neurone de commande, dont l’activation suffit à déclencher l’évasion, mais la situation est probablement un peu plus complexe.
En parallèle, les cellules LC4 et LPLC2 réalisent des connections avec d’autres neurones descendants (12). C’est l’activation simultanée d’autres DNs qui produirait les réponses en mode-long, on encore les marches arrière. On retrouverait donc des circuits parallèles, l’un dédié aux urgences (par un neurone de « commande ») et l’autre aux réponses avec plus de préparations (par une population de neurones).
Rappelons également que nous voyons le monde avec une fréquence de 60Hz, alors les mouches voient le monde avec une fréquence de rafraîchissement d’environ 250Hz (elles voient plus d’images par seconde). C’est l’une des raisons pour laquelle il est si difficile d’attraper une mouche.
Figure 5.1 : Evasion chez la Drosophile (a) basée sur un looming stimulus visuel.
(a) Envol chez la drosophile en réponse à un prédateur. Le comportement d’évasion peut être sub-divisé en sous-comportements qui peuvent être interrompus. On distingue une évasion longue, qui inclut une élévation des ailes, rendant le vol post-décollage plus stable, et une évasion courte, pour les urgences, qui supprime l’étape d’élévation des ailes, gagnant plusieurs précieuses millisecondes, mais nécessitant des manœuvres aériennes pour corriger la trajectoire après coup – (a1, b1, c1) Notion de looming stimulus. Lorsqu’un prédateur s’approche, cela produit une « tâche » dont la taille grossit au cours de l’approche d’un prédateur – (c2) La taille angulaire θ(t) de cette tâche augmente de manière non linéaire au cours du temps, et ne dépend que du ratio L/v – (d) corrélation linéaire positive entre l’imminence du looming (plus le temps l/v est petit, plus la collision arrive rapidement), et la proportion d’évasion courtes chez la Drosophile -D’après (7) (Source : P. Clémençon)
B2. Chez le criquet
En présence d’un prédateur approchant, les criquets peuvent réaliser une réponse d’évasion composée de sous-comportements qui peuvent être interrompus : flexion initiale, co-contraction et saut (figure 5.2). La biomécanique est assez remarquable car les pattes réalisent une accélération de 100g lors de la détente !
Figure 5.2 : Evasion chez le criquet (b) basée sur un looming stimulus visuel. Le saut chez le criquet peut être divisé en 3 sous-comportements qui peuvent être stoppés : flexion initiale, co-contraction des muscles flexeurs et extenseurs puis saut (Source : P. Clémençon)
Les neurones clés sont le Descending Contralateral Movement Detector (DCMD) et le lobula giant movement detector (LGMD1 et LGMD2).
Le LGMD, situé dans le lobula, est connecté avec le DCMD, un neurone descendant. Le DCMD est un neurone avec un axone de large diamètre (15 um) qui est connecté avec les motoneurones thoraciques, déclenchant l’évasion (flexion initiale, co-contraction des muscles flexeurs et extenseurs, puis détente). Chaque potentiel d’action enregistré dans le DCMD correspond exactement à un potentiel d’action émis par le LGMD. Autrement dit, pour enregistrer l’activité du LGMD, on peut enregistrer à la place l’activité du DCMD (« plus simple ») et vice-versa.
Le LGMD (et donc le DCMD) réagit sélectivement à des objets approchant (looming stimuli) (13). indépendamment de leur forme (rond ou carré), texture (damier ou motif plein), ou de la température. Dans une expérience ayant conduit à l’obtention d’un prix IgNobel, Rind & Simmons ont enregistré l’activité du LGMD en projetant le film Star Wars à un criquet. Ils ont pu corréler les pics d’activité du LGMD avec les moments où un vaisseau s’approche de la caméra.
Le LGMD possède un large arbre dendritique, divisé en 3 sous-régions : 2 branches mineures et l’arbre majeur, qui ressemble à un éventail et qui reçoit des informations de cellules sensibles à la direction du mouvement. On observe expérimentalement que le pic d’activité du LGMD (fréquence maximale de potentiels d’action) coïncide avec le moment où la taille angulaire ϴ(t) du prédateur dépasse un seuil ϴseuil, avec un petit délai fixe (14)
En termes algorithmiques, le LGMD se comporte comme un détecteur de seuil angulaire, et réalise une multiplication (combinaison de L et 1/v) dont l’implémentation n’est pas entièrement comprise (15). D’autres neurones sensibles au looming ont été découverts, et un traitement de l’information pourrait avoir lieu en parallèle du LGMD.
Ainsi, on retrouve des similarités et des différences dans l’organisation anatomique du système visuel des criquets et Drosophiles ainsi que dans l’algorithme de détection du looming (extraction du mouvement, extraction des caractéristiques de looming, opération de multiplication, et action) (figure 6).
Figure 6 : Circuits sous-jacent l’évasion chez la Drosophile et le criquet. L’information visuelle provenant des photorécepteurs est pré-traitée ensuite par les neurones de la lamina et de la médulla, pour extraire notamment la direction du mouvement (EMD). En aval, les neurones LPLC2, LC4 (drosophile) et le LGMD (criquet) calculent les informations pertinentes pour la détection de collisions imminentes. Chez la drosophile et le criquet, Le DCMD (criquet) et la Fibre Géante (drosophile) sont connectés à des motoneurones des ganglions thoraciques et ‘’ commandent ‘’ certains aspects de l’évasion, en fonction du rapport taille/vitesse l/v calculé.
Le schéma du circuit de la Drosophile est directement inspiré de (33). Celui du criquet est inspiré de (34),et insiste plus sur les calculs réalisés, l’identité des cellules étant moins connue que chez la Drosophile (Source : P. Clémençon)
C) Système auditif
L’écholocation, utilisée par les chauve-souris pour localiser leurs proies, est également leur talon d’Achille. De nombreux insectes volants nocturnes possèdent des « oreilles », une fine membrane de cuticule appelée tympan couplée à des organes chordotonaux. Ces tympans sont également associés avec des sacs aériens. Il existe une grande diversité de forme et de placement de ces oreilles (figure 7a) (17).
Figure 7 : Perception et réponse comportementale aux ultrasons chez les insectes – a) localisation des oreilles chez divers insectes – b) localisation du tympan chez une mante religieuse – c) Chronophotographie du plongeon abrupt d’une mante P. agrionina lors de la présentation d’ultrasons (flèche blanche « cri de chauve-souris » sur la figure) (16). Les zooms avant et après la présentation du cri de chauve-souris montrent les ajustements posturaux complexes (Source : P. Clémençon)
La vibration du tympan lors de la réception d’un son conduit à l’excitation de neurones sensoriels (A1 et A2 chez les papillons de nuit). L’influx nerveux est traité par les interneurones des ganglions thoraciques (lien) (réduction du bruit et sommation temporelle du signal). Ensuite, des motoneurones sont alors activés et coordonnent une réponse d’évasion, comme un plongeon d’urgence (figure 7). Chez de nombreux papillons, le seuil d’activation des cellules A1 et A2 est bas pour des fréquences autour de 20kHz, fréquences utilisées par la sérotine brune (figures 8a & 8b).
L’attaque d’une chauve-souris commence par de simples clics, et, si un papillon est choisi comme cible (l’attaque est alors imminente), la fréquence de clics augmente : c’est la phase de buzz terminal. Comme présenté sur les enregistrements électrophysiologiques (4) de la figure 8c soulignent, les cellules A1 (rouge) réagissent à des sons de faibles intensité, permettant de détecter les chauve-souris lorsque la phase d’approche commence à peine. L’asymétrie entre les réponses des cellules A1 situés à gauche ou à droite du papillon l’informe de la localisation de la chauve-souris. Il est intéressant de noter que les neurones aux seuils les plus bas peuvent détecter les cris des chauve-souris à un moment où l’écho retour est trop faible pour être audible pour les chauve-souris, donnant un avantage crucial pour le papillon. Les cellules A2 ont un seuil d’intensité plus haut et leur activation indiquerait un danger imminent, comme le buzz terminal d’une chauve-souris.
Figure 8 : Audition et évasion chez les insectes, avec une emphase sur le système proie-prédateur chauve-souris/noctuelle – a) Traitement des ultrasons chez les papillons de nuit : structure du tympan et cellules sensorielles associées – b) courbe de sensibilité de neurones A1 en fonction de la fréquence testée chez différentes espèces de papillons de nuit – c) enregistrements électrophysiologiques extracellulaires chez un papillon de nuit en présence d’un stimulus mimant l’attaque d’une chauve souris (en gris) L’attaque d’une chauve-souris se décompose en une phase de recherche, une phase d’approche (c1) et une phase de buzz terminal durant laquelle la fréquence des cris augmente (c2) – Enregistrements électrophysiologiques et courbes de sensibilités tirés de (4) (Source : P. Clémençon)
D) Système de perception des flux d’air
Les blattes, les grillons et plusieurs Hexapodes « basaux » (thysanoures, lépismes, …) (lien) possèdent à l’arrière de l’abdomen des cerques (figure 9) (18), un système sensoriel généraliste impliqué dans de nombreux comportements : reproduction, perception de la gravité et fuite notamment. Ces cerques sont recouverts de poils mécanosensoriels couplés chacun à un neurone. Lorsque l’inclinaison du poil dépasse un certain seuil, le neurone couplé produit un influx nerveux (lien).
Le système est à ce jour le plus sensible du règne animal, 1/10ème de l’énergie d’un photon suffirait à activer les neurones mécanosensoriels (19).
Les poils sont de taille variable, entre 0.2 et 2 mm. Une des interprétations fonctionnelles proposées est la suivante : lorsqu’un prédateur arrive à toute vitesse, les poils courts réagiraient avant les poils longs, et le recrutement successif des poils indiqueraient un danger imminent. Lorsqu’un prédateur arrive lentement, les poils courts ne collecteraient pas assez d’énergie pour dépasser l’angle seuil à l’inverse des poils longs qui indiqueraient la présence du prédateur (20) (lien). Les poils présentent également un angle préférentiel. Le neurone couplé à un poil sélectif aux déplacements d’air transversaux réagira plus fortement à un prédateur venant du côté qu’un prédateur venant de l’arrière. L’hétérogénéité dans la canopée de poils refléterait la large gamme de flux d’air qu’un grillon pourrait rencontrer dans son environnement.
L’information provenant de tous ces poils est traitée dans le ganglion abdominal terminal (TAG) composé d’une vingtaine d’interneurones géants permettent une vitesse de conduction rapide, et d’une trentaine de neurones locaux. Les propriétés sélectives des poils permettraient d’effectuer un multiplexing d’information. Par exemple, les synapses des poils présentant le même angle préférentiel se situent à peu près au même endroit dans le ganglion abdominal terminal, et cela coïncide avec les dendrites (inputs) de 4 interneurones du TAG présentant également des directions préférées (figure 9). Ces 4 interneurones répondent de manière maximale à des stimulus provenant de 45, 135, 225, 315°. Ces 4 neurones formeraient une unité fonctionnelle suffisante pour encoder la direction du stimulus. (21)
Concernant l’identité du flux d’air, les grillons distingueraient un prédateur et le vent ambiant ou les propres mouvements de l’animal via l’encodage du profil d’accélération. Le LGI, le plus gros interneurone, est l’un des candidats.
On dit souvent que l’information provenant de 1 000 poils est compressée dans un système comprenant moins d’une centaine de neurones. En fait, il ne s’agit pas d’une compression, mais plutôt d’une extraction des caractéristiques les plus pertinentes : l’identité, la direction et l’intensité du flux d’air. Lorsqu’un journal enroulé se dirige vers une blatte, l’information pertinente est qu’un objet dangereux arrive vite, et non le titre du journal.
Figure 9 : Perception des flux d’air et évasion chez le grillon. Le déplacement d’air d’une araignée est obtenu par Particle Image Velocimetry (22). Il provoque la déflexion des poils filiformes, qui sont couplés à des neurones. L’information provenant des poils est traitée dans le TAG, et des interneurones géants envoient l’information vers des motoneurones dans les ganglions thoraciques. La cartographie en bas à droite souligne que les synapses des poils possédant la même préférence angulaire se situent au même endroit dans le TAG (21). Dessin de ganglion inspiré de (23) (Source : P. Clémençon)
E) Intégration multimodale et aspects cognitifs
D’autres systèmes de détection non décrits ici des prédateurs incluent : le toucher et la perception des vibrations (par des sensilles campaniformes, organes chordonotaux et subgénuaux) ou l’odorat (par des sensilles olfactives).
L’évasion est multisensorielle et les différentes modalités décrites précédemment interagissent. Le cerveau combine ces entrées sensorielles variées, en les pondérant, pour ne produire qu’une sortie comportementale. Une modalité sensorielle peut induire une augmentation de la sensibilité d’une autre. Par exemple, chez les mâles de noctuelle Spodoptera, la sensibilité des neurones olfactifs à la phéromone (olfaction) augmente quelques heures après l’exposition de cris de chauve-souris (audition) (24).
Les insectes peuvent réaliser des apprentissages non associatifs (habituation, sensibilisation, …) et associatifs (conditionnement). Lors d’attaques répétées, on peut observer une sensibilisation se traduisant par un état d’alerte prolongé, une diminution du temps de réaction, une augmentation de la FID (distance proie-prédateur nécessaire pour que la proie fuit) ou encore des changements de stratégies. A l’inverse, l’exposition répétée d’un stimulus non nocif peut conduire à des phénomènes d’habituation comme chez les larves de moustiques (25) (26).
Concernant les apprentissages associatifs, les insectes peuvent apprendre les zones de prédations (par exemple les fourmis et les pièges des fourmillons), et certains insectes, même non sociaux comme par exemple les grillons des bois (27), apprennent un danger en apprenant de conspécifiques (social learning).
L’évasion est le résultat de l’intégration et pondération de multiples modalités sensorielles ainsi que de connaissances, ne produisant au final qu’un seul output comportemental, une grande question en neurosciences.
III) L’évasion des insectes et l’Homme
Nous discuterons de deux points : l’apport de la compréhension des mécanismes d’évasion pour anticiper certaines réponses aux changements globaux, et l’utilisation des connaissances sur les circuits neuraux sous-jacents à l’évasion pour la bio-inspiration (figure 10).
A) Biomimétisme
L’évasion peut se résumer en un « calcul sensorimoteur donnant un comportement flexible dans un temps contraint ».
Pour les aspects sensoriels, les structures impliqués dans l’évasion ont parfois donné lieu à des implémentations technologiques (figure 10). La compréhension des mécanismes physiques de la déflexion des poils filiformes du grillon a inspiré le développement de micro-senseurs de flux d’air, des MEMS, qui pourraient être utilisés pour détecter d’éventuelles fuites, invisibles à l’œil nu (28). Les pattes sauteuses puissantes du criquet inspirent des actionneurs mécaniques capable de stocker de l’énergie élastique et de la libérer rapidement.
Pour les aspects neuronaux, les circuits sous-jacents l’évasion intéressent de près les ingénieur.e.s en neuromorphique (29) et en robotique. La contrainte de rapidité se traduit probablement par un nombre limité de connections synaptiques, et une simplicité structurale en comparaison à d’autres circuits. Le système visuel du criquet a par exemple inspiré plusieurs algorithmes de détection des collisions (30), qui ont été implémentés dans des puces électroniques (31), ou dans des robots (9).
Pour les aspects plus comportementaux, la biomécanique spectaculaire de certains systèmes, comme les pattes des criquets, inspirent des systèmes dits de ressort. Certaines sociétés utilisent les connaissances sur l’évasion chez les papillons de nuit pour concevoir des drones destinés à être déployés dans les serres pour éliminer les papillons de nuit nuisibles aux récoltes, sans affecter les autres insectes.
Figure 10: Boucle sensori-motrice de l’évasion : implications technologiques et menaces liées aux changements globaux (Source : P. Clémençon)
B) Changements globaux
Les modifications de température et le bruit anthropique pourrait altérer la performance musculaire et sensorielle des insectes comme décrit chez les poissons (32) (figure 10). Cette diminution des capacités pourrait avoir des conséquences en chaîne sur les écosystèmes (33) (34). Brièvement, plusieurs études soulignent la probable plus grande sensibilité des prédateurs aux températures par rapport à leurs proies (35), ainsi que l’augmentation potentielle de prédation intra-guilde du fait de la modification des niches (36) (37), ce qui pourrait conduire à terme à une prolifération d’herbivores nuisibles dans les récoltes. L’arrivée de nouveaux prédateurs par dispersion anthropique, comme le frelon asiatique, peut avoir de larges conséquences, les proies ne possédant pas de systèmes sensorimoteurs adaptés (« matched », voir partie I) à ces prédateurs.
Concernant l’évasion en elle-même, plusieurs altérations des réponses d’évasion ont été observées.
Les syrphes qui utilisent le mimétisme batésien comme défense anti-prédateur primaire pourraient voir leur taux de prédation augmenter suite à des changements de phénologie (38).
Certains polluants, même en capacités sublétales, peuvent altérer les capacités d’évasion en diminuant les capacités d’habituation ainsi que l’activité spontanée des neurones du système visuel du criquet (39). La pollution lumineuse, comme les lampes aux vapeurs de mercure (certes beaucoup moins utilisées aujourd’hui), inhibent la perception des ultrasons par les papillons, et donc diminuent leur capacité à évader aux chauve-souris (40). Une augmentation du CO2 atmosphérique pourrait quant à elle diminuer les capacités d’évasion, comme cela a été montré chez les pucerons (41). Des perturbations du développement pourrait se traduire par des altérations des structures sensorielles et motrices, diminuant les capacités évasives des insectes.
L’extinction de certaines espèces d’insectes se traduit également par une perte irrémédiable de savoirs fondamentaux potentiels (en neurosciences, sciences évolutives, écologie,…) qui auraient pu par exemple être transposées de manière plus appliquée (dans des technologies comme décrit plus haut par exemple).
Chaque espèce raconte une histoire particulière (principe de Krogh : « en sélectionnant le bon modèle animal, on peut maximiser le ratio signal sur bruit, rendant l’interprétation des résultats beaucoup plus simple (42), et la connaissance des mécanismes ne peut se faire qu’avec des organismes dits « modèles ».
Conclusion
L’évasion chez les insectes offre un cadre de travail combinant terrain et de laboratoire. L’élicitation répétable des réponses d’évasion et le faible nombre de neurones impliqués (du fait de la parcimonie neurale liés aux contraintes de vitesse) sont des avantages pratiques.
La flexibilité dans les réponse offre néanmoins la possibilité d’étudier comment le systèmes nerveux extraient l’information et prennent une décision motrice appropriée dans un temps contraint.
Les fortes pressions anthropiques peuvent se traduire en des opportunités manquées de mieux comprendre les systèmes proies-prédateurs. La diversité des innovations évolutives pour échapper aux prédateurs apporte des connaissances fondamentales mais également appliquées en inspirant par exemple de nouvelles technologies.
Paul Clémençon
Je suis fasciné par les insectes depuis très longtemps, et plus particulièrement par les algorithmes computationnels sous-jacents leur perception du monde. J’ai eu l’opportunité de réaliser plusieurs stages sur des modèles insectes (nervation des ailes chez les Hyménoptères, écologie sensorielle de la migration du papillon bogong, perception olfactive d’une noctuelle dans un environnement bruité, …).
Je suis actuellement doctorant à l’Institut de Recherche sur la Biologie de l’Insecte (IRBI) où j’étudie principalement la mécano-réception chez le grillon, en combinant des modèles de réseaux de neurones et théorie des jeux, et des approches expérimentales (électrophysiologie et comportement).
Un grand merci à Jérôme Casas et Sylvia Anton pour leur relecture !
Bibliographie
D. A. Evans ; A. V. Stempel ; R. Vale & T. Branco (2019) : Cognitive Control of Escape Behaviour. Trends Cogn. Sci. 23, 334–348 (lien)
D. D. Yager (2010) : “Predator Evasion” in Encyclopedia of Animal Behavior, M. D. Breed, J. Moore, Eds. (Academic Press, 2010), pp. 765–773
R. Wehner (1978) : « Matched filters » — neural models of the external world. J. Comp. Physiol. A161, 511–531 (lien)
J. H. Fullard ; J. W. Dawson & D. S. Jacobs (2003) : Auditory encoding during the last moment of a moth’s life. J. Exp. Biol. 206, 281–294 (lien)
J. H. Fullard, et al. (2012) : Release from bats: genetic distance and sensoribehavioural regression in the Pacific field cricket, Teleogryllus oceanicus. Naturwissenschaften97, 53–61 (lien)
G. M. Card (2012) : Escape behaviors in insects. Curr. Opin. Neurobiol.22, 180–186 (lien)
C. R. von Reyn, et al. (2017) : Feature Integration Drives Probabilistic Behavior in the Drosophila Escape Response. Neuron 94, 1190-1204.e6 (lien)
H. S. Cheong ; I. Siwanowicz & G. M. Card (2020) : Multi-regional circuits underlying visually guided decision-making in Drosophila. Curr. Opin. Neurobiol.65, 77–87 (lien)
S. Bermúdez I Badia ; U. Bernardet & P. F. M. J. Verschure (2010) : Non-Linear Neuronal Responses as an Emergent Property of Afferent Networks: A Case Study of the Locust Lobula Giant Movement Detector. PLOS Comput. Biol.6, e1000701 (lien)
M. Y. Peek & G. M. Card (2016) : Comparative approaches to escape. Curr. Opin. Neurobiol.41, 167–173 (lien)
J. M. Ache, et al. (2019) : Neural Basis for Looming Size and Velocity Encoding in the Drosophila Giant Fiber Escape Pathway. Curr. Biol.29, 1073-1081.e4 (lien)
M. Dombrovski, et al.(2023) : Synaptic gradients transform object location to action. Nature613, 534–542 (lien)
F. C. Rind & P. J. Simmons (1992) : Orthopteran DCMD neuron: a reevaluation of responses to moving objects. I. Selective responses to approaching objects. J. Neurophysiol.68, 1654–1666 (lien)
F. Gabbiani, et al.(2004) : Multiplication and stimulus invariance in a looming-sensitive neuron. J. Physiol.-Paris98, 19–34 (lien)
F. Gabbiani ; H. G. Krapp ; C. Koch & G. Laurent (2002) : Multiplicative computation in a visual neuron sensitive to looming. Nature420, 320–324 (lien)
D. D. Yager ; M. L. May & M. B. Fenton (1990) : Ultrasound-triggered, flight-gated evasive maneuvers in the praying mantis Parasphendale agrionina. I. Free flight. J. Exp. Biol.152, 17–39 (lien)
D. D. Yager (1999) : Structure, development, and evolution of insect auditory systems. Microsc. Res. Tech.47, 380–400 (lien)
Y. Baba & H. Ogawa (2017) : “Cercal System-Mediated Antipredator Behaviors” in The Cricket as a Model Organism: Development, Regeneration, and Behavior, H. W. Horch, T. Mito, A. Popadić, H. Ohuchi, S. Noji, Eds. (Springer Japan, 2017), pp. 211–228 (lien)
T. Shimozawa ; J. Murakami & T. Kumagai (2003) : “Cricket Wind Receptors: Thermal Noise for the Highest Sensitivity Known” in Sensors and Sensing in Biology and Engineering, F. G. Barth, J. A. C. Humphrey, T. W. Secomb, Eds. (Springer, 2003), pp. 145–157 (lien)
T. Steinmann & J. Casas (2017) : The morphological heterogeneity of cricket flow-sensing hairs conveys the complex flow signature of predator attacks. J. R. Soc. Interface14, 20170324 (lien)
G. A. Jacobs ; J. P. Miller & Z. Aldworth (2008) : Computational mechanisms of mechanosensory processing in the cricket. J. Exp. Biol.211, 1819–1828 (lien)
J. Casas ; T. Steinmann & O. Dangles (2008) : The Aerodynamic Signature of Running Spiders. PLOS ONE3, e2116 (lien)
D. Booth ; B. Marie ; P. Domenici ; J. M. Blagburn & J. P. Bacon (2009) : Transcriptional Control of Behavior: Engrailed Knock-Out Changes Cockroach Escape Trajectories. J. Neurosci.29, 7181–7190 (lien)
S. Anton ; K. Evengaard ; R. B. Barrozo ; P. Anderson & N. Skals (2011) : Brief predator sound exposure elicits behavioral and neuronal long-term sensitization in the olfactory system of an insect. Proc. Natl. Acad. Sci.108, 3401–3405 (lien)
N. F. Zilber-Gachelin & M. P. Chartier (1973) : Modification of the Motor Reflex Responses Due to Repetition of the Peripheral Stimulus in the Cockroach: I. Habituation at the Level of an Isolated Abdominal Ganglion. J. Exp. Biol. 59, 359–381 (lien)
H. Baglan ; C. Lazzari & F. Guerrieri (2017) : Learning in mosquito larvae (Aedes aegypti): Habituation to a visual danger signal. J. Insect Physiol.98, 160–166 (lien)
I. Coolen ; O. Dangles ; J. Casas (2005) : Social Learning in Noncolonial Insects? Curr. Biol.15, 1931–1935 (lien)
G. J. M. Krijnen ; H. Droogendijk ; A. M. K. Dagamseh ; R. K. Jaganatharaja & J. Casas (2014) : “Crickets as Bio-Inspiration for MEMS-Based Flow-Sensing” in Flow Sensing in Air and Water: Behavioral, Neural and Engineering Principles of Operation, H. Bleckmann, J. Mogdans, S. L. Coombs, Eds. (Springer, 2014), pp. 459–488 (lien)
T. Dalgaty ; E. Vianello ; B. De Salvo & J. Casas (2018) : Insect-inspired neuromorphic computing. Curr. Opin. Insect Sci.30, 59–66 (lien)
S. Yue ; F. C. Rind ; M. S. Keil ; J. Cuadri & R. Stafford (2006) : A bio-inspired visual collision detection mechanism for cars: Optimisation of a model of a locust neuron to a novel environment. Neurocomputing69, 1591–1598 (lien)
D. Jayachandran, et al.(2020) : A low-power biomimetic collision detector based on an in-memory molybdenum disulfide photodetector. Nat. Electron.3, 646–655 (lien)
P. Domenici ; B. J. M. Allan ; C. Lefrançois & M. I. McCormick (2019) : The effect of climate change on the escape kinematics and performance of fishes: implications for future predator–prey interactions. Conserv. Physiol.7, coz078 (lien)
J. A. Harvey (2015) : Conserving host–parasitoid interactions in a warming world. Curr. Opin. Insect Sci.12, 79–85 (lien)
A. N. Laws (2017) : Climate change effects on predator–prey interactions. Curr. Opin. Insect Sci.23, 28–34 (lien)
M. E. Moore ; C. A. Hill & J. G. Kingsolver (2021) : Differing thermal sensitivities in a host–parasitoid interaction: High, fluctuating developmental temperatures produce dead wasps and giant caterpillars. Funct. Ecol.35, 675–685 (lien)
M. Pélissié ; F. Johansson & C. Hyseni (2002) : Pushed Northward by Climate Change: Range Shifts With a Chance of Co-occurrence Reshuffling in the Forecast for Northern European Odonates. Environ. Entomol.51, 910–921 (lien)
B. T. Barton & O. J. Schmitz (2009) : Experimental warming transforms multiple predator effects in a grassland food web. Ecol. Lett.12, 1317–1325 (lien)
C. Hassall ; J. Billington & T. N. Sherratt (2019) : Climate-induced phenological shifts in a Batesian mimicry complex. Proc. Natl. Acad. Sci.116, 929–933 (lien)
R. H. Parkinson ; S. Zhang & J. R. Gray (2020) : Neonicotinoid and sulfoximine pesticides differentially impair insect escape behavior and motion detection. Proc. Natl. Acad. Sci.117, 5510–5515 (lien)
A. M. Svensson & J. Rydell (1998) : Mercury vapour lamps interfere with the bat defence of tympanate moths (Operophtera spp.; Geometridae). Anim. Behav.55, 223–226 (lien)
W. T. Hentley ; A. J. Vanbergen ; R. S. Hails ; T. H. Jones & S. N. Johnson (2014) : Elevated Atmospheric CO2 Impairs Aphid Escape Responses to Predators and Conspecific Alarm Signals. J. Chem. Ecol.40, 1110–1114 (lien)
S. Lindstedt & Krogh 1929 or ‘The Krogh Principle.’ J. Exp. Biol.217, 1640–1641 (2014)
K. T. Sillar ; L. D. Picton & W. J. Heitler (2016) : The Neuroethology of Predation and Escape (John Wiley & Sons, 2016).
Les interactions proies-prédateurs sont répandues dans la culture populaire, et fascinent par la grande diversité d’innovations évolutives dans la course aux armements proies-prédateurs. Certaines font l’objet de plusieurs documentaires spectaculaires sur la BBC, et d’autres ont inspiré de nombreuses scènes de films et jeux vidéos.
Les insectes, pour leurs avantages expérimentaux, offrent une bonne porte d’entrée pour l’étude des interactions proies-prédateurs ainsi que les bases neurales du comportement d’évasion. Les réponses d’évasion sont facilement élicitables, robustes, soumises à des contraintes de parcimonie neuronale, mais ne sont pas figées pour autant (1). Elles offrent un cadre scientifique permettant d’étudier des questions comme le decision-making ou la sélection d’action parmi un répertoire d’actions comportementales. L’évasion offre également la possibilité d’avoir une riche approche comparative, c’est-à-dire comprendre comment un même problème est résolu par différentes espèces dans des niches écologiques variées, de la détection et l’identification de la menace au choix de l’action comportementale appropriée.
La compréhension des mécanismes d’évasion inspire les ingénieurs pour développer de nouvelles technologies, et est cruciale pour prédire l’évolution des relations proies-prédateurs suite aux pressions anthropiques.
I – Écologie des jeux de cache-cache et de course-poursuite entre prédateurs et proies
Dans cet article, nous nous focaliserons plus sur l’échelle de l’individu que sur l’échelle macroécologique. Lorsque l’on parle d’interactions proies-prédateur, on pense souvent en premier lieu aux aspects de course-poursuite (défenses secondaires), mais il ne faut pas oublier les lignes de défenses primaires en amont. Les lignes de défenses, primaires ou secondaires, peuvent être actives ou passives (nécessitent ou non la présence de comportement « actif »).
Les interactions proies-prédateurs peuvent être décomposées en deux parties (figure 1): une phase de cache-cache, et une phase de course-poursuite. Cette séquence peut ensuite se répéter (2). Un exemple illustratif est décrit en détail par Fabre : lorsqu’une araignée est attaquée par une guêpe maçonne, elle tombe au sol (course-poursuite), échappant quelque temps à la prédation. La guêpe inspecte attentivement les alentours (cache-cache) avant de retrouver l’araignée. Cette dernière court (course-poursuite), pour échapper à la guêpe, et la séquence peut se répéter plusieurs fois (3).
Figure 1 : Cadre conceptuel pour analyser les interactions proies-prédateurs, en fonction de la connaissance que les protagonistes ont de la présence de l’autre. L’embuscade, c’est-à-dire quand le prédateur est « conscient » de la présence de la proie avant celle-ci, ne sera pas abordée ici. Division de l’évènement de prédation basée sur (4) et quelques défenses anti-prédateurs associées à chaque étape (Source : P. Clémençon)
A) Cache-cache
Dans un jeu de cache-cache entre proie et prédateur, on modélise souvent la proie comme un agent cherchant à minimiser la probabilité de détection. Différentes stratégies minimisent cette probabilité (5). On peut citer notamment la sélection d’habitats complexes et difficiles d’accès pour le prédateur (galles, refuges), des phénologies particulières, l’évitement des zones de prédation (les grillons percevant l’odeur d’araignées changent de zone) (6) ou encore l’homochromie, parfois combinée à des couleurs voyantes. Les criquets Oedipoda sont par exemple bien camouflés dans l’environnement, leur corps prenant un ton ocre rappelant la couleur des sentiers de cailloux.
De plus, leurs ailes postérieures (visibles seulement lorsque l’insecte saute) présentent des couleurs vives. Cela augmente l’efficacité du camouflage, le prédateur devant changer d’objet visuel à chercher, et cela limite la persistance des attaques en limitant la probabilité de détection.
Dans le cas précis où la proie perçoit la présence du prédateur, mais où le prédateur ne perçoit pas la présence de la proie (jeu de cryptisme), certaines modifications du comportement ont été décrites. On peut citer l’arrêt du chant des Orthoptères (figure 2a) (7), le repositionnement derrière une feuille (figure 2b), le vol erratique des papillons de nuit (figure 2c), certains mouvements pour augmenter l’efficacité du camouflage (oscillations chez les phasmes) ou des comportements d’immobilisation et d’alerte qui diminuent la probabilité de détection (figure 2d) et augmentent l’attention.
Il faut noter que la détection du prédateur n’entraîne pas automatiquement une réponse de fuite chez la proie. L’évasion présente de nombreux coûts en terme de fitness : réduction du temps passé à s’approvisionner ou à rechercher un partenaire. Certains modèles soulignent également la perte de l’effet de surprise ainsi que d’avantages tactiques si la proie s’enfuit trop tôt (8).
Figure 2 : Quelques comportements observés pour minimiser la probabilité de détection de la proie par le prédateur – (a) Arrêt du chant et mouvements d’antennes chez un grillon – (b) repositionnement – (c) vol erratique, rendant la localisation difficile pour les chauve-souris – (d) immobilisation et préparation au saut. Notons que la préparation du saut peut être avortée si la menace disparaît (Source : P. Clémençon)
Lorsque la proie est détectée, le prédateur ne lance pas systématiquement une attaque, la proie pouvant dissuader le prédateur.
Certains signaux, structures ou comportements indiquent un danger. Les mantes présentent une posture menaçante (figure 3b), les Apocrites présentent un aiguillon, parfois couplé à une glande à venin, d’autres insectes présentent des épines ou autres structures de défense, plusieurs papillons ont des motifs en forme « yeux » pour effrayer le prédateur, … Certains insectes présentent des signaux aposématiques (couleurs vives, clics par les organes tympanaux chez les papillons de nuit destinés aux chauve-souris, figure 3a) qui indiquent un mauvais goût (9). Lorsque plusieurs espèces toxiques présentent les mêmes signaux (comme chez les papillons Heliconius), on parle de mimétisme mullérien. A l’inverse, certaines espèces inoffensives imitent des espèces toxiques, on parle de mimétisme batésien.
D’autres structures, signaux et comportements rendent la proie difficile à attaquer (les papillons-lunes présentent des queues sur les ailes postérieures qui interfèrent avec le système de détection des chauves souris (10)) ou peu attractive (de nombreux insectes réalisent une thanatose c’est-à-dire faire le mort (figure 1).
Encart : Mentionnons quelques bénéfices de la stratégie de groupe. L’existence de comportements de vigilance permet de s’approvisionner plus efficacement, un ou plusieurs conspécifiques donnant l’alerte (many-eyes hypothesis). Le groupe dilue également le risque de capture (en perturbant le prédateur qui peine à trouver une cible, et en diminuant le risque d’être pris pour cible), et certains comportements collectifs rebutent le prédateur (shimmering waves chez certaines abeilles) (11).
B) Course-poursuite
Lorsque le prédateur lance une attaque, on parle de jeu différentiel. Quand toutes les barrières de défenses « primaires » décrites au dessus ont échoué, une réponse active « secondaire » de la proie est requise pour éviter d’être mangée. La détermination du « gagnant » dépend de composants variés : aspects sensoriels, performance locomotrice et trajectoires. On retrouve chez les insectes des circuits spécialisés, efficaces et rapides minimisant le délai de traitement et d’initiation de la réponse d’évasion (voir Partie II), et parfois des mécanismes biomécaniques puissants, comme les pattes arrière de certains Orthoptères.
Figure 3 : (a) Dissuasion : (a1) émission active de signaux aposématiques (clics chez un papillon Arctidé peu appétant). (a2) posture de défense chez une mante religieuse.
(b) Contre-attaque : (b1) Interférence avec le buzz terminal de la chauve souris par émission de signaux sonores chez le papillon B.trigona – (b2) coup de patte défensif chez le grillon Acheta face à une guêpe parasitoïde Liris. (b3) défense chimique chez le bombardier Brachinus sp. (source inconnue)
(c) Fuite : (c1) chute du puceron en réponse à la phéromone d’alarme – (c2) plongée d’une larve de moustique – (c3) rotation du corps et course chez la blatte Periplaneta en réponse à un prédateur (indiqué par la flèche jaune). La blatte tourne selon un angle choisi aléatoirement parmi plusieurs directions possibles (indiquées par des triangles rouges) dans un même secteur angulaire. Adapté de (12) (c4) saut chez le criquet – (c5) envol chez la Drosophile avec ou sans élévation des ailes, en fonction de l’urgence – (c6) Manœuvres ou chute chez un insecte nocturne volant en réponse aux cris d’une chauve-souris – (c7) évasion post-ingestion chez le coléoptère R. attenuata. D’après Shinji Sugiura, Kobe University – (c8) fuite vers un terrier chez le grillon des champs (Source : P. Clémençon)
B.1) Diversité du répertoire comportemental pour échapper aux prédateurs
Durant la phase de subjugation (figure 1), les réponses d’évasion sont variées (13).
Certains insectes émettent des signaux ou réalisent des comportements pour étourdir le prédateur. Plusieurs espèces de grillons et de blattes utilisent l’« attaque » et se défendent des guêpes parasitoïdes en donnant des coups de pattes puissants (14) (figure 3b2). Certains insectes produisent une substance lorsqu’ils sont attaqués : les pucerons produisent une substance collante qui aveugle les coccinelles, et les bombardiers libèrent une substance brûlante (figure 3b2). Si plusieurs papillons de nuit émettent de simples clics aposématiques comme décrit précédemment (15) (figure 3a1), les noctuelles Bertholdia trigona produisent des signaux plus complexes qui couvrent une large gamme de fréquences et perturbent l’écholocation des chauve-souris (brouillage sonore ou sonar jamming), le timing de l’écho retour étant crucial pour détecter avec précision la position d’un objet dans l’espace (16). Les chauves-souris manquent les papillons B. trigona de 15-20 cm (figure 3b1).
D’autres insectes font de l’autotomie (perte volontaire d’un membre) (17) ou présentent une fausse tête, perturbant le prédateur, ce qui donne quelques millisecondes cruciales à la proie pour s’échapper.
Enfin, de nombreuses proies réalisent une réponse de fuite, c’est-à-dire un déplacement pour sortir de la « zone de danger», en sortant du champ de détection du prédateur par des manœuvres diverses (Partie B.2) : chute vers le sol (figure 3c1), plongée en spirale (figure 3c2), course (figure 3c3), rotation du corps suivi d’un mouvement de translation comme le saut (figure 3c4), vol (figures 3c5 & c6), nage dans le tube digestif (figure 3c7). La présence de refuges influence l’évasion (figure 3c8).
Course-poursuite d’une Araignée Pardosa et un grillon Nemobius : le grillon essaie de sortir de la zone de danger – ralentie 2 fois (Source : Jérôme Casas)
Il faut noter que les réponses d’évasion ne sont pas fixes, comme illustré sur la figure 4. Le comportement d’envol d’une drosophile peut être décomposé en plusieurs sous-comportements (ajustements posturaux, élévation des ailes, saut) qui peuvent être interrompus ou sautés en fonction de la nature de la menace. De même, chez le criquet, la flexion initiale ne s’ensuit pas toujours de l’étape de co-contraction, et cette étape ne s’ensuit pas toujours d’un saut (on parle de réponse abortive).
B.2) Les trajectoires d’évasion, un mélange entre optimum géométrique, imprédictibilité et recherche de refuges
Plusieurs paramètres géométriques sont couramment utilisés pour caractériser les trajectoires d’évasion : l’angle de rotation initial du corps, la sinuosité de la trajectoire, la distance totale parcourue (Distance Fled), ou encore le timing (FID pour Flight Initiation Distance). De nombreux modèles géométriques permettent de prédire les trajectoires et le timing en fonction de différents paramètres.
Le ratio entre la vitesse du prédateur et celle de la proie (18), ou celui entre les distances d’attaques et de fuite (19) ont souvent été décrits comme des paramètres clés dans la compréhension des interactions proies-prédateurs.
Dans certains cas, une action locomotrice (un saut, un cycle de marche) suffit. Par exemple, dans la litière de feuilles, lorsqu’une araignée Pardosa attaque un grillon des bois Nemobius, celui-ci peut réaliser un saut puissant qui l’éloigne parfois à plusieurs dizaines de centimètres de l’araignée, et sort de la zone de danger (pour l’araignée, les indices sensoriels indiquant la position du grillon dans l’environnement ne sont alors plus suffisants pour lancer une attaque) (20).
Vidéo d’une attaque d’araignée Pardosa sur un grillon des bois Nemobius (ralentie environ 300 fois)
A l’inverse, dans le cas où une chauve-souris poursuit un papillon de nuit, le ratio des vitesses est en faveur du prédateur (21). Les trajectoires de fuite résultant en une évasion réussie sont souvent caractérisées par une forte accélération angulaire, la présence de zigzaging, ou l’exécution de plongeons d’évasion en spirales imprédictibles qui rend leur tracking sensoriel plus difficile pour les chauve-souris (figure 3c5).
Vitesse et manœuvres sont des déterminants du succès de l’évasion
La persistance du prédateur influence fortement le comportement de fuite. Chez les criquets ou les larves de moustiques, la distance parcourue augmente lors d’attaques répétées. L’angle de fuite est caractérisé par un mélange entre optimum géométrique et imprédictibilité. On constate intuitivement l’angle de fuite qui maximise la distance au prédateur est généralement de 180°. Certains écarts ont été interprétés comme étant la conséquence de bruit dans la perception et dans la réponse motrice, ou encore de contraintes locomotrices.
Les punaises Triatoma (Reduviidae), par exemple, fuient à un angle de 120°, interprété comme un angle maximisant la distance au prédateur tout en gardant sa position dans le champ visuel (22). D’autres insectes fuient assez clairement dans des directions aléatoires, imprédictibles, une stratégie limitant l’apprentissage chez les prédateurs, en particulier dans le cas d’attaques persistantes. L’imprédictibilité souligne la flexibilité des réponses d’évasion, qui sont plus complexes que de simples boucles réflexes comme cela a été décrit dans le passé.
Choisir un angle totalement aléatoire (comportement protéen) présente des limites, la proie pouvant foncer vers le prédateur. Chez les blattes, des études ont mis en évidence la présence de plusieurs directions préférées dans un même secteur angulaire, choisies aléatoirement, qui ne sont néanmoins pas dirigées vers le prédateur (angles de 90° – 270°) (12).
Ainsi, l’imprédicitibilité (randomness) est aussi un déterminant du succès de l’évasion, en limitant l’apprentissage.
Enfin, des expériences ont souligné l’orientation de certaines trajectoires vers des refuges. De nombreux insectes sur les parties aériennes des végétaux se laissent simplement tomber au sol lorsqu’un prédateur approche. L’un des exemples les plus célèbres est celui des pucerons qui se laissent tomber sur le sol lorsqu’il sentent l’haleine d’un herbivore, ou lorsqu’il perçoivent la phéromone d’alarme émise par un conspécifique, le β-farnesène (figure 3c1). Certains insectes construisent un refuge, comme les grillons des champs, et s’y cachent lorsqu’un prédateur s’approche de trop près pendant une durée qui dépasse généralement la patience du prédateur (23). D’autres insectes fuient vers une cachette de proximité comme la végétation ou une pierre. L’entrée dans un refuge n’est pas sans coûts en terme de fitness. Les larves et pupes de moustiques quittent la surface pour fuir la prédation en surface, mais leurs réserves d’oxygène diminuent (24).
Ainsi, la connaissance de routes ou la présence de refuges influencent la trajectoire, même si les refuges ne sont pas sans coûts.
Pour conclure, rappelons que les réponses d’évasion ne sont pas de simples boucles réflexes. Elles dépendent également de nombreux facteurs internes et externes comme l’âge, l’expérience, la taille corporelle, la température ou encore le comportement en cours.
Les décisions prises lors de l’évasions sont le résultat d’une évaluation des coûts/bénéfices (figure 4) (25). Par exemple, un papillon de nuit mâle très proche d’une femelle ne s’enfuit que lorsqu’une chauve-souris est sur le point d’attaquer (buzz terminal). Au contraire, ce même papillon de nuit loin d’une femelle réalise des réponses d’évasion dès que le prédateur est détecté, lors de la phase d’approche (26).
Figure 4 : Aspects multifactoriels de l’évasion : de nombreux facteurs internes et externes influencent les décisions prises lors de l’évasion (Source : P. Clémençon)
Le point de vue du prédateur
Il existe un gradient de stratégies chez les prédateurs à différentes échelles.
Pour la rencontre proie-prédateur, d’un côté du spectre se trouvent les prédateurs sit-and-wait, qui attendent qu’une proie passe à proximité comme les chenilles carnivores, certaines libellules restent immobiles sur une perche (lien), un point d’observation. Certains construisent des pièges comme larves de fourmillons, ou les toiles. D’autres produisent des signaux trompeurs pour attirer la proie jusqu’à eux (araignée Bolas, guêpes sur toile,…). De l’autre côté du spectre se trouvent les prédateurs cruising (27) qui parcourent activement des distances jusqu’à trouver une proie, comme certaines coccinelles.
La recherche de proies commence souvent par la détection d’indices à longue portée (CO2, …). Une fois la proie détectée, les prédateurs sit-and-wait attendent le dernier moment pour attaquer (la distance entre la proie et le prédateur est très petite, et le temps de réaction de la proie est souvent supérieur à la durée de l’interaction). Certains prédateurs, comme les mantes religieuses réalisent une embuscade lente avant l’assaut final rapide. D’autres prédateurs initient, une fois la proie détectée, une course-poursuite effrénée. Les cicindèles, asilides et libellules sont connus pour leur capacité à traquer leur proie à grande vitesse. On distingue deux stratégies de poursuite, le suivi, ou tracking, et la navigation parallèle qui consiste à viser un point fictif où la proie se retrouvera à un instant t+1 (lien) (28).
Bien entendu, certains prédateurs utilisent diverses stratégies en fonction de facteurs internes et externes. La distance d’attaque des mantes religieuses Tenodera dépend du niveau de satiété : une mante affamée va parcourir de grandes distances pour se rapprocher de la proie, alors qu’une mante repue adopte des comportements plus opportunistes, n’attaquant que les proies à faible portée (29).
Tout comme la proie, le succès du prédateur repose sur des systèmes sensoriels efficaces mais aussi des biomécaniques puissants et précis, notamment des systèmes de catapulte. L’exemple le plus frappant est celui de la larve de libellule, qui a inspiré le film Alien, un système rapide et précis (lien).
Ne manquez pas la seconde partie de cet article sur les aspects plus neuroéthologiques de l’évasion !Partie II
Paul Clémençon
Je suis fasciné par les insectes depuis très longtemps, et plus particulièrement par les algorithmes computationnels sous-jacents leur perception du monde. J’ai eu l’opportunité de réaliser plusieurs stages sur des modèles insectes (nervation des ailes chez les Hyménoptères, écologie sensorielle de la migration du papillon bogong, perception olfactive d’une noctuelle dans un environnement bruité, …).
Je suis actuellement doctorant à l’Institut de Recherche sur la Biologie de l’Insecte (IRBI) où j’étudie principalement la mécano-réception chez le grillon, en combinant des modèles de réseaux de neurones et théorie des jeux, et des approches expérimentales (électrophysiologie et comportement).
Un grand merci à Jérôme Casas et Sylvia Anton pour leur relecture !
Bibliographie
G. M. Card (2012) : Escape behaviors in insects. Curr. Opin. Neurobiol.22, 180–186 (lien)
S. Alpern & S. Gal (2006) : The Theory of Search Games and Rendezvous (Springer Science & Business Media, 2006) (lien)
J. Fabre :Souvenirs entomologiques: Etudes sur l’instinct et les murs des insectes, vol. 4 (1891)
J. A. Endler : “Interactions between predator and prey” in Krebs, J.R. and Davies, N., Behavioural Ecology, Blackwell Scientific Publications, pp. 169–196
W. J. Boeing : “Defensive Avoidance” in Encyclopedia of Animal Behavior, M. D. Breed, J. Moore, Eds. (Academic Press, 2010), pp. 476–480
R. Kortet & A. Hedrick (2004) : Detection of the Spider Predator, Hololena nedra by Naïve Juvenile Field Crickets (Gryllus integer) Using Indirect Cues. Behaviour141, 1189–1196 (lien)
A. V. Hedrick (2000) : Crickets with extravagant mating songs compensate for predation risk with extra caution. Proc. R. Soc. Lond. B Biol. Sci.267, 671–675 (lien)
M. Broom (2013) : “Interactions Between Searching Predators and Hidden Prey” in Search Theory: A Game Theoretic Perspective, S. Alpern, R. Fokkink, L. Gąsieniec, R. Lindelauf, V. S. Subrahmanian, Eds. (Springer, 2013), pp. 233–248
G. D. Ruxton ; W. L. Allen ; T. N. Sherratt & M. P. Speed (2023) : Avoiding Attack: The Evolutionary Ecology of Crypsis, Aposematism, and Mimicry (Oxford University Press, 2018) https:/doi.org/10.1093/oso/9780199688678.001.0001
J. R. Barber, et al. (2015) : Moth tails divert bat attack: Evolution of acoustic deflection. Proc. Natl. Acad. Sci.112, 2812–2816 (lien)
G. Kastberger ; F. Weihmann & T. Hoetzl (2010) : Complex social waves of giant honeybees provoked by a dummy wasp support the special-agent hypothesis. Commun. Integr. Biol.3, 179–180 (lien)
P. Domenici ; J. M. Blagburn & J. P. Bacon (2011) : Animal escapology I: theoretical issues and emerging trends in escape trajectories. J. Exp. Biol.214, 2463–2473 (lien)
P. W. Bateman & P. A. Fleming (1990) : “Invertebrates” in, pp. 177–196
R. Hustert & W. Gnatzy (1995) : The Motor Program for Defensive Kicking in Crickets: Performance and Neural Control. J. Exp. Biol.198, 1275–1283 (lien)
A. J. Corcoran ; J. R. Barber ; N. I. Hristov & W. E. Conner (2011) : How do tiger moths jam bat sonar? J. Exp. Biol.214, 2416–2425 (lien)
A. J. Corcoran & W. E. Conner (2012) : Sonar jamming in the field: effectiveness and behavior of a unique prey defense. J. Exp. Biol.215, 4278–4287 (lien)
P. A. Fleming ; D. Muller & P. W. Bateman (2007) : Leave it all behind: a taxonomic perspective of autotomy in invertebrates. Biol. Rev.82, 481–510 (lien)
P. Domenici & G. D. Ruxton (2015) : “Prey behaviors during fleeing: escape trajectories, signaling, and sensory defenses” in Escaping From Predators: An Integrative View of Escape Decisions, D. T. Blumstein, J. Cooper William E., Eds. (Cambridge University Press, 2015), pp. 199–224
J. Casas & T. Steinmann (2014) : Predator-induced flow disturbances alert prey, from the onset of an attack. Proc. R. Soc. B Biol. Sci.281, 20141083 (lien)
S. Morice ; S. Pincebourde ; F. Darboux ; W. Kaiser & J. Casas (2013) : Predator-prey pursuit-evasion games in structurally complex environments. Integr. Comp. Biol.53, 767–779 (lien)
A. J. Corcoran & W. E. Conner (2016) : How moths escape bats: predicting outcomes of predator–prey interactions. J. Exp. Biol.219, 2704–2715 (lien)
C. Lazzari & D. Varjú (1990) : Visual lateral fixation and tracking in the haematophagous bug Triatoma infestans. J. Comp. Physiol. A167, 527–531 (lien)
D. M. Hugie (2003) : The waiting game: a “battle of waits” between predator and prey. Behav. Ecol.14, 807–817 (lien)
I. Rodríguez-Prieto ; E. Fernández-Juricic & J. Martín (2006) : Anti-Predator Behavioral Responses of Mosquito Pupae to Aerial Predation Risk. J. Insect Behav.19, 373–381 (lien)
R. C. Ydenberg & L. M. Dill (1986) : “The Economics of Fleeing from Predators” in Advances in the Study of Behavior, (Elsevier, 1986), pp. 229–249.
N. Skals ; P. Anderson ; M. Kanneworff ; C. Löfstedt & A. Surlykke (2005) : Her odours make him deaf: crossmodal modulation of olfaction and hearing in a male moth. J. Exp. Biol.208, 595–601 (lien)
N. Zoroa ; M. J. Fernández-Sáez & P. Zoroa (2011) : A foraging problem: Sit-and-wait versus active predation. Eur. J. Oper. Res.208, 131–141 (lien)
S. T. Fabian ; M. E. Sumner ; T. J. Wardill ; S. Rossoni & P. T. Gonzalez-Bellido (2018) : Interception by two predatory fly species is explained by a proportional navigation feedback controller. J. R. Soc. Interface15, 20180466 (lien)
S. C. Pickard, et al.(2021) : Internal state effects on behavioral shifts in freely behaving praying mantises (Tenodera sinensis). PLOS Comput. Biol.17, e1009618 (lien)
Cerveau de drosophile illustré par imagerie électronique (Source : Kei Ito et al.)
Que ce soit chez les arthropodes ou les vertébrés, le cerveau constitue le principal centre de contrôle de l’organisme. Chez les insectes, le cerveau reçoit des informations sensorielles (influx nerveux) provenant à la fois des organes des sens de la tête (vision, olfaction et goût par exemple) via les interneurones ascendants, et des ganglions plus postérieurs du thorax et de l’abdomen (voir article sur l’anatomie du système nerveux chez les insectes :lien). Bien que composé en grande partie des corps cellulaires d’interneurones et de fibres nerveuses, il comprend également les motoneurones (neurones moteurs) des muscles antennaires.
Certains de ces interneurones s’étendent le long du cordon nerveux, jusqu’aux ganglions les plus postérieurs (extrémité de l’abdomen), pour transmettre des informations comme celles en lien avec la coordination des mouvements du corps.
Le cerveau des insectes est subdivisé en trois régions : le protocérébron, le deutocérébron et le tritocérébron (figure 1).
Figure 1 : Représentation d’un cerveau de Locusta – (a) Vue antérieure – (b) Vue latérale (d’après Albrecht, 1953) (Source : Insects – Structure and function – Chapman Ed5) (Modifié par B. GILLES)
Le protocérébron constitue la partie la plus volumineuse du cerveau et la plus importante au plan fonctionnel. Situé dans la partie antérieure de la tête, de forme bilobée, il s’étend latéralement avec les lobes optiques (centres photorécepteurs des ocelles et des yeux composés – lien) et comprend les corps pédonculés et le complexe central, deux zones particulièrement développées chez les insectes sociaux (figures 2 et 3).
Le protocérébron joue un rôle primordial dans les sécrétions endocrines (molécules sécrétées directement dans l’hémolymphe), un rôle comparable à celui de l’hypothalamus chez les vertébrés. Les cellules neurosécrétrices, dont le nombre peut dépasser la centaine, sont le plus souvent regroupées dans un axe antéro-dorsal au niveau d’un système neuroendocrinien central appelé la « pars intercerebralis » (figures 2 et 3) (§ suivant). Cette région intervient notamment dans la régulation des cycles éveil-sommeil (neurones octopaminergiques).
D’autres cellules neurosécrétrices sont quant à elles reliées à un autre système de forme allongée : le corps cardiaque (également appelé : corpora cardiata ou corpus cardiacum) qui, comme l’indique ce nom, est accolé à l’aorte contractile située à l’arrière du cerveau (système assurant la circulation de l’hémolymphe). Le corps cardiaque influe à la fois sur la mue, la maturation ovocytaire, la pigmentation et les mouvements péristaltiques des tubes de Malpighi et du tube digestif au niveau du proctodeum (lien), ainsi que sur les hormones émises par les cellules neurosécrétrices de la pars intercerebralis .
Figure 2 : Présentation des principales régions de neuropiles organisés (ombré) et des corps cellulaires (points noirs) d’un cerveau d’insecte (Source : Insects – Structure and function – Chapman Ed5) (Modifié par B. GILLES)Figure 3 : Représentation 3D d’un cerveau de Manduca sexta femelle (Sphingidae) en vue antérieure (A), dorsale (B) et postérieure (C) – Barre d’échelle 500 µm (Source : El Jundi et al., 2009)
Corps pédonculés
Situés de chaque côté de la pars intercerebralis (figures 2 et 3), les corps pédonculés, interviennent dans l’intégration des signaux sensoriels comme l’apprentissage, la mémoire ou la discrimination olfactive.
Chacun d’eux est constitué d’un calice dans sa partie antérieure à partir duquel s’étend ventralement un pédoncule se divisant en deux lobes, parfois trois, appelés α, β, et γ (figures 2 et 4a). La terminologie peut prêter à confusion puisque les deux lobes principaux sont parfois dits lobes ventral et médian selon leur orientation.
Le calice, pouvant être simple ou double, se subdivise en 3 sous-unités (neuropiles) connus sous le nom de lèvre, collet et anneau basal (figure 4b).
Regroupant les neurones olfactifs, les calices sont doubles chez des espèces à l’odorat développé comme les abeilles Apis et les blattes Periplaneta, et simples, voire absents, chez des espèces où l’olfaction joue au contraire un rôle mineur comme chez les libellules et les cigales.
Chose étonnante, le calice est chez certaines espèces associé à la gustation et est de petite taille.
Figure 4 : Exemples de neurones associés aux corps pédonculés (a) Neurones intrinsèques (cellules de Kenyon – numérotées 1, 2 et 3) montrant différents types d’arborisation dendritique dans le calice. Notez que l’axone du neurone 1 ne s’étend que dans le lobe β ; pour le neurone 2, seule la partie proximale est représentée ; l’axone du neurone 3 s’étend à la fois aux lobes ⍺ et β (Apis) (d’après Mobbs, 1985) – (b) Neurone d’entrée du lobe antennaire. L’arborisation dendritique est limitée à un seul glomérule, mais ce n’est pas le cas de tous les neurones d’entrée du lobe antennaire aux corps pédonculés. L’axone a des branches vers les deux parties du calice (Manduca) (basé sur Homberg, 1994) (Source : Insects – Structure and function – Chapman Ed5) (Modifié par B. GILLES)
Les relations du calice avec l’olfaction étant présentes chez la plupart des taxons, il est suggéré qu’il s’agisse d’un caractère ancestral (plésiomorphie). Ces relations et l’organisation de l’innervation du calice sont toutefois fortement soumises à l’influence de l’environnement et des comportements des espèces avec des spécialisations évolutives uniques. Par exemple, les punaises de la famille des Notonectidae, de par leur mode de vie subaquatique, possèdent des antennes et des lobes antennaires atrophiés.
La taille relative des corps pédonculés est corrélée à la complexité du comportement des insectes : petits chez les collemboles (lien), les Hétéroptères, les Diptères et les Odonates, de taille moyenne chez les Coléoptères, les Orthoptères, les Blattodae, les Lépidoptères et les Tenthrèdes (Hyménoptères symphytes), et plus développés chez les insectes sociaux (figure 5a). Par exemple, chez les abeilles et guêpes sociales dont les comportements de chasse reposent en grande partie sur l’acuité visuelle (informations visuelles provenant des lobes optiques), les calices sont imposants ce qui est probablement lié au rôle important que joue la vision dans l’apprentissage de zones d’alimentation, des sites de nidification et même dans l’identification des congénères de la colonie.
Image de neurones dans un cerveau d’abeille domestique (Source : Stanley Heinze et al.)
Les espèces de Coléoptères phytophages généralistes (plusieurs types de plantes hôtes) possèdent par exemple des pédoncules de taille supérieure à celles ayant des régimes plus spécialisés. Les criquets pèlerins en phase grégaire, ayant des régimes alimentaires diversifiés et des interactions sociales complexes, ont également des corps pédonculés plus volumineux que les criquets en phase solitaire. Chez les termites, où le volume du pédoncule et des lobes a et b est important, le calice ne représente qu’environ un tiers du volume total des corps pédonculés. Chez les abeilles et les guêpes sociales, il représente près 70% du volume des corps pédonculés.
La partie antérieure des corps pédonculés regroupe un grand nombre d’interneurones, les cellules de Kenyon. Bien qu’il ait été inventorié une diversité morphologique et fonctionnelle de ces cellules entre les taxons, la structure type d’une cellule de Kenyon demeure stable entre les taxons. Elles sont constituées d’un corps cellulaire situé au-dessus du calice, de dendrites (prolongements ramifiés des neurones) s’insérant dans le calice, d’un axone courant le long du pédoncule se ramifiant ventralement dans les lobes α et β (figure 4a).
Les abeilles ouvrières possèdent environ 170 000 cellules de Kenyon dans chaque pédoncule, représentant 40% de tous les neurones du cerveau ; la mouche Calliphora possède quant à elle environ 21 000 cellules de Kenyon dans chaque pédoncule, soit environ 12% de la totalité de ces neurones.
Figure 5 : (a) Les volumes des corps pédonculés sont exprimés par rapport à la taille du complexe central. Les corps pédonculés sont relativement plus volumineux chez les espèces sociales que chez les autres. Notez que chez les termites (Isoptera), le pédoncule et les lobes représentent la majeure partie du volume, alors que chez les Hymenoptera il s’agit du calice (d’après Howse, 1974) – (b) Modifications du nombre de fibres dans les corps pédonculés de Drosophila. Les insectes représentés par les histogrammes étaient tous âgés de 21 jours après l’éclosion. Les environnements offrant moins de stimulation ont entraîné un plus petit nombre de fibres chez les mâles et les femelles (d’après Technau, 1984) – (c) Variations de volume des différentes parties du corps pédonculés chez Apis. Le volume des différentes parties augmente à mesure que les insectes vieillissent. Les butineuses ont des lèvres et des colliers plus gros dans les calices que les nourrices du même âge et de leurs parents (d’après Durst et al., 1994) (Source : Insects – Structure and function – Chapman Ed5) (Modifié par B. GILLES)
Il a été démontré que des changements se produisaient dans les corps pédonculés avec l’âge et l’expérience. Chez la drosophile par exemple, le volume des corps pédonculés augmente de plus de 20% durant la première semaine de la vie d’adulte (figure 5b), un phénomène également observé durant les dix premiers jours de la vie des abeilles ouvrières (figure 5c). Bien que certaines espèces de grillons et de Coléoptères aient la capacité de produire de nouvelles cellules de Kenyon au cours de leur vie d’adulte, cette faculté disparaît chez d’autres lorsque l’insecte atteint l’âge adulte ou durant le stade nymphal.
L’augmentation du volume corporel des corps pédonculés résulte de l’augmentation du nombre de fibres mais aussi de l’expérience vécue par l’insecte. Chez la drosophile et l’abeille Apis, par exemple, le nombre de fibres présents dans les pédoncules augmente davantage chez les individus ayant des contacts avec leurs congénères que chez des individus isolés. De la même manière, chez les abeilles ouvrières, les corps pédonculés sont plus imposants chez celles qui butinent que chez les nourrices du même âge (figures 5b et 5c).
Les corps pédonculés sont impliqués dans l’apprentissage olfactif et l’apprentissage visuel chez certains insectes. Des études suggèrent qu’ils pourraient intervenir dans certains processus de prise de décision. Des drosophiles mutantes présentant des malformations au niveau des corps pédonculés ont montré un comportement globalement normal mais avec une incapacité à apprendre certains types d’associations ou des comportements stéréotypés et certaines difficultés à répondre aux nouvelles conditions.
Exemples de la diversité des cerveaux d’insectes adultes (a) Drosophila melanogaster – (b) Tribolium castaneum – (c) Apis mellifera – (d) Schistocerca gregaria – (Basé sur Rein et al. 2002 ; Kurylas et al. 2008 ; Dreyer et al. 2010 ; Rybak et al. 2010). Tous les cerveaux ont été dimensionnés à la même largeur et les neuropiles respectifs ont le même code de couleur : lobes antennaires (bleu), corps pédonculés (rouge), lamina des lobes optiques (jaune), lobula des lobes optiques (orange), complexe central (vert) (Source : Koniszewski et al., 2016)
Complexe central
Le complexe central est une zone riche en interneurones où se rencontrent quatre neuropiles formant 16 colonnes (8 de chaque côté du cerveau) de neurones empilés en couches distinctes (figures 6a, b et c).
Les fonctions possibles du complexe central se précisent lentement. Bien que les neurones le composant aient été identifiés comme répondants aux stimuli mécanosensoriels, visuels et chimiques, et comme déclenchant des schémas comportementaux (neurones moteurs), le cheminement de l’information entrant et sortant du complexe central est encore loin d’être entièrement établi. Le rôle le plus caractérisé du complexe central serait le traitement des informations visuelles de la région du bord dorsal de l’œil composé qui est sensible à la lumière polarisée.
Figure 6 : Exemples de neurones du complexe central (criquet du genre Schistocerca) (d’après Homberg, 1991, 1994) – (a) Neurone reliant le pont protocérébral à la partie inférieure du corps central – (b) Deux neurones reliant les lobes accessoires latéraux à des parties du corps central – ( c ) Deux neurones symétriques avec une arborisation étendue dans le corps central supérieur (Source : Insects – Structure and function – Chapman Ed5) (Modifié par Benoît GILLES)
Les insectes utilisent le plan de polarisation du ciel (lien) (plan variant au cours de la journée en fonction de la position du soleil) pour s’orienter et se repérer dans l’espace même par temps nuageux. Ces informations visuelles permettent à l’insecte de disposer d’une sorte de boussole céleste.
Il est également admis que le complexe central intervient dans la surveillance et l’organisation des schémas comportementaux en lien avec la locomotion. Des enregistrements de l’activité électrique des neurones de cette zone chez des cafards, à l’aide de réseaux multi-électrodes, ont montré que leur activité était corrélée au rythme de la marche et que, dans de nombreux cas, le changement d’activité des neurones précédait le changement de comportement. De plus, la stimulation électrique de ces neurones à travers les électrodes pouvait modifier les habitudes de marche. D’autres travaux, dont plusieurs études utilisant des mutants de drosophiles, ont suggéré un rôle général du complexe central dans l’intégration sensorielle motrice (propriorécepteurs) en particulier en relation avec la position du corps et différentes régions du corps par rapport à l’espace extérieur.
Lobes optiques
Les lobes optiques forment des extensions latérales reliant le protocérébron et les yeux composés. Chacun d’eux est constitué de trois neuropiles successifs (figures 2 et 3), connus sous le nom de : lamina, medulla et lobula.
Les arrangements structuraux en couches des neurones offrent un fort degré de rétinotopie, c’est-à-dire que la représentation précise de toute image arrivant sur l’œil (cellules photoréceptrices) est conservée à mesure que le traitement de l’information nerveuse progresse de la lamina au lobula. De manière étonnante, les fibres se croisent horizontalement entre les neuropiles successifs, formant des chiasmes optiques externe et interne. Cette configuration nerveuse entraine des inversions successives de la carte neurale de l’image visuelle le long de l’axe antéro-postérieur (figures 2, 3 et 7).
Figure 7 : Représentation des interneurones à champ large et à champ étroit avec leurs arborisations dans la lobula et leurs connexions avec les interneurones descendants chez la mouche (d’après Gronenberg et Strausfeld, 1992) (Source : Insects – Structure and function – Chapman Ed5) (Modifié par B. GILLES)
Deutocérébron
Le deutocérébron, essentiellement associé aux antennes, est constitué de deux lobes antennaires regroupant à la fois les neurones du système olfactif (chémorécepteurs) et ceux des centres mécanosensoriels et moteurs (figures 1 et 2).
Ces régions, relativement mal définies, contiennent les arborisations terminales des neurones mécanosensoriels provenant du scape et du pédicelle, voire également du flagelle, de l’antenne ainsi que les arborisations dendritiques des motoneurones contrôlant les muscles antennaires.
La détection des sources de nourriture, des sites de pontes et des partenaires sexuels repose en grande partie sur la chimioréception, le contact, l’olfaction ou la gustation (lien). La répartition sur les antennes et les pièces buccales des récepteurs olfactifs, gustatifs et de contact est spécifique à chaque espèce.
Les lobes antennaires, constituées de neuropiles synaptiques denses, sont appelés glomérules, leur nombre variant considérablement d’une espèce à l’autre (figure 8a). Par exemple, la blatte Periplaneta en compte 125, le papillon Sphingidae du genre Manduca environ 64 et le moustique Aedes moins de 10.
De manière générale, les axones des sensilles olfactives des antenne se terminent dans les glomérules, où chacun d’eux n’est relié qu’à un seul glomérule (figure 8b). Chez les criquets, possèdant plus de 1 000 glomérules mal définis, chaque axone innerve a contrario plusieurs glomérules.
Au sein d’une même espèce, les glomérules semblent conserver une forme et une position constantes. Les mâles d’espèces détectant leurs partenaires sexuelles à l’aide des phéromones émises par les femelles possèdent cependant deux ou trois glomérules de plus grande taille, formant le complexe macroglomérulaire.
L’ensemble des informations nerveuses provenant des chémorécepteurs antennaires exprimant les mêmes protéines réceptrices olfactives convergent vers un même glomérule, se connectent au niveau des synapses (zone de contact entre deux neurones) avec des interneurones (neurones de projection) puis transmettent l’information aux calices des corps pédonculés et à des zones spécifiques du protocerebrum (figures 4b, 8b).
Figure 8 : (a) Représentation de glomérules dans un lobe antennaire – (b) Représentation d’un interneurone local et d’un neurone de sortie (de projection) (d’après Matsumoto et Hildebrand, 1981) – (c) Un neurone sérotoninergique avec des terminaisons dans tous les glomérules et une ramification étendue à travers d’autres parties du cerveau (d’après Kent et al., 1987) (Source : Insects – Structure and function – Chapman Ed5) (Modifié par B. GILLES)
Les cellules chimioréceptrices des antennes ne s’activant qu’en présence d’un composé volatile (COV : composé organique volatil) en lien avec l’écologie de l’espèce (phéromone sexuelle par exemple) ou d’un mélange de composés spécifiques (sources alimentaires par exemple), cela permet de créer une transmission linéaire et directe avec le glomérule où les informations sont véhiculées sans aucune ambiguïté. En effet, le nombre relativement important de glomérules et d’interneurones permet aux insectes de distinguer un large panel d’odeurs.
Des neurones de projection réagissent toutefois lorsque l’antenne est exposée à une large gamme d’odeurs fournissant ainsi à l’insecte des informations générales sur son environnement.
On retrouve également dans les lobes antennaires une deuxième population d’interneurones : les neurones locaux. Contrairement aux neurones de projection, ces neurones s’interconnectent à plusieurs glomérules (figure 8b). Ils diffèrent également considérablement des neurones de projection par leur morphologie, leurs schémas de connectivité, leur physiologie et leurs neurotransmetteurs. Les abeilles possèdent par exemple environ 4 000 interneurones locaux dans chaque lobe antennaire.
Les interneurones locaux interviennent dans des voies nerveuses soit inhibitrice, soit excitatrice. Les neurones locaux inhibiteurs sécrètent des neurotransmetteurs comme l’acide aminobutyrique (GABA) et l’histamine, les excitateurs l’acétylcholine.
Il a été mis en évidence que des interneurones des lobes antennaires rejoignaient d’autres zones du cerveau. Ces interneurones synthétisent plusieurs types de neurotransmetteurs comme la sérotonine (neurone sérotoninergique), la dopamine, l’octopamine et/ou des peptides dont les profils (proportions et nombre de molécules) diffèrent considérablement selon les espèces d’insectes. Il semblerait que ces interneurones interviennent dans l’association des odeurs à des qualités gustatives appétissantes ou aversives (figure 7c).
Tritocérébron
Le tritocérébron, situé sous le deutocérébron, est à la fois la plus petite région du cerveau et la moins étudiée. Il est relié au ganglion sous-oesophagien par une paire de lobes enserrant le tube digestif et interconnectés par une commissure post-oesophagienne parfois peu visible (figure 2). Dans sa partie antérieure, le tritocérébron reçoit les informations nerveuses provenant des neurones sensoriels et moteurs du labrum et des pièces buccales (lien).
Comme le deutocérébron, le tritocérébron donne naissance à des neurones descendants vers les ganglions thoraciques et abdominaux. Bien qu’une certaine architecture générale ait pu être décrite, peu de choses sont connues quant à ses relations avec les autres régions du cerveau ou quant à ses connexions intrinsèques.
Illustration 3D du système nerveux d’une abeille (Source : Eric Keller – Bioopatone)
The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
El Jundi B. ; Huetteroth W. ; Kurylas A.E. & Schachtner J. (2009) : Anisometric brain dimorphism revisited : implementation of a volumetric 3D standard brain in Manduca sexta. The Journal of Comparative Neurology, 517,2,210-225 (lien)
Fahrbach S.E. (2006) : Structure of the mushroom bodies of the insect brain. Annu. Rev. Entomologiste. 51:209-32 (lien)
Koniszewski N.D.B. et al. (2016) : The insect central complex as model for heterochronic brain development – background, concepts and tools. Development Genes and Evolution, 226, 209-219 (lien)
Pfeiffer K. & Homberg U. (2014) : Organization and functional roles of the central complex in the insect brain. Annual review of entomology. 59:165-184 (lien)
Identique à celui des autres arthropodes, le système nerveux des insectes, est constitué d’une chaîne nerveuse composée de ganglions reliés entre eux par des cordons nerveux. Ces ganglions regroupent les corps cellulaires (également appelé soma ou péricaryon) de l’ensemble des interneurones et des motoneurones ainsi que les extensions dendritiques et axoniques des neurones (encadré en bas de page). Les axones des motoneurones s’étendent des nerfs périphériques tandis que les axones des neurones sensoriels se dirigent vers les ganglions.
La structuration des ganglions comprend une zone périphérique constituée des corps cellulaires, formant le cortex, et une zone centrale où s’arborisent les neurones qui s’y insèrent (site d’intégration synaptique), appelée la neuropile. Au sein de cette masse de fibres nerveuses, certaines fibres possèdent une orientation commune antéro-postérieure (connectifs), d’autres une orientation transversale reliant la moitié gauche et droite du ganglion (commissures).
Primitivement, chaque segment de l’insecte comportait un ganglion responsable du traitement des informations sensorielles locales provenant des récepteurs musculaires des membres et de la paroi corporelle. Cet arrangement demeure, aujourd’hui, partiellement visible chez de nombreux embryons d’insectes, structure qui se réorganise durant le développement post-embryonnaire. Certains ganglions segmentaires fusionnent avec leurs voisins pour former des neuromères. Le cerveau des insectes correspond à la fusion de plusieurs ganglions de la région céphalique.
Le système nerveux central est représenté par le cerveau, positionné dorsalement au-dessus de l’intestin antérieur (position dite préorale), et d’une série de ganglions segmentaires situés ventralement.
Dans un axe antéro-postérieur, le premier ganglion de la chaîne ventrale faisant suite au cerveau est, comme son nom l’indique, le ganglion sous-œsophagien. Il résulte de la fusion des neuromères des segments mandibulaire, maxillaire et labial. Chez certains insectes, en particulier les Hyménoptères et les Diptères, le ganglion sous-œsophagien est lui-même fusionné au cerveau, ne laissant qu’un petit conduit à travers lequel passe l’œsophage. Ce ganglion innerve les pièces buccales (lien) (mandibules, maxillaires et labium), les muscles du cou et les glandes salivaires.
Dans le thorax, il y a généralement trois ganglions. Toutefois, chez certains insectes, deux ou trois d’entre eux sont fusionnés comme chez la mouche Musca (figures 1a et 1b). Le ganglion métathoracique est fusionné avec au moins le premier neuromère abdominal chez presque tous les Ptérygotes (insectes ailés). De chaque côté des ganglions thoraciques diffusent cinq ou six nerfs se ramifiant pour innerver les muscles et les sensilles du thorax et de ses appendices (ailes et pattes) (lien).
Figure 1a : Nombre de ganglions ventraux (d’après Horridge, 1965). (a) La plupart des ganglions restent séparés (Dictyopterus, Coleoptera) – (b) Tous les ganglions ventraux, à l’exception du sous-œsophagien, sont fusionnés en une seule masse ganglionnaire composée (Musca, Diptera) (Source : Structure and fonction – Chapman – Ed5-2013)
Dans l’abdomen, le plus grand nombre de ganglions abdominaux rencontrés, tant chez des larves et des adultes insectes, est de 8. C’est le cas par exemple chez les Thysanoures ou le mâle Pulex (Siphonaptères). Le dernier ganglion abdominal dérive quant à lui de la fusion des quatre derniers segments abdominaux. Bien que plus nombreux, les ganglions abdominaux sont plus petits et diffusent moins de nerfs que ceux du thorax. De plus, il est constaté une ramification moins importante et diversifiée des nerfs dans l’abdomen. Ces caractéristiques découlent du fait que l’abdomen abrite peu d’organes liés à la motilité nécessitant la présence de neurones moteurs (motoneurones), mais davantage d’organes liés à la digestion et à la reproduction.
Chez certaines espèces, l’ensemble des ganglions abdominaux sont fusionnés comme chez la punaise hématophage Rhodnius et les mouches Cyclorrhapha (en référence à l’ouverture circulaire par laquelle l’adulte sort de la pupe) (figure 2a). En règle générale, un processus de fusion des ganglions se déroule entre la phase larvaire et adulte des espèces holométaboles (lien). Le degré de fusion peut être très variable entre les taxons, même proches phylogénétiquement, entre les mâles et les femelles et selon le statut social chez les abeilles sans dard Melipone.
Figure 1b : illustration 3D du système nerveux d’une abeille (Source : Eric Keller – Bioopatone)
Dans la plupart des cas, les muscles d’un segment sont innervés par des neurones du ganglion du même segment, mais une certaine innervation par des axones provenant des ganglions voisins se produit également (figure 2). Certaines fibres afférentes peuvent également être inter-segmentaires.
Figure 2 : Micrographie électronique d’une jonction muscle/nerf – Mise en évidence de la zone de contact entre la terminaison atonale et la membrane musculaire (d’après Smith et Treherne, 1963) (Source : Structure and fonction – Chapman – Ed5-2013)
Neurones du système nerveux central
Neurones moteurs
Les motoneurones interviennent dans le contrôle des muscles. Leur nombre, relativement variable, dépend du nombre d’unités musculaires que contient le segment. Chaque muscle est innervé par 2 à 11 motoneurones. Par exemple, le ganglion métathoracique de la blatte Periplaneta contient 500 neurones.
Chez les insectes, les corps cellulaires des motoneurones sont relativement de grande taille. Par exemple, dans les ganglions métathoraciques de criquets, leur diamètre varie de 20µm à 90µm. Au sein des ganglions, la structuration des motoneurones est symétrique (homologie droite et gauche) et demeure constante entre les individus. Le neurite de chaque motoneurone (prolongement du corps cellulaire d’un neurone : axone ou dendrite) augmente de diamètre en entrant dans la neuropile, au sein de laquelle il donne naissance à une ramification arborescente (figure 3). La forme de cet arbre dendritique est caractéristique de chaque motoneurone, et sa complexité est une indication de celle de ses connexions synaptiques (connexion entre deux neurones) avec les autres neurones. Contrairement aux vertébrés, les insectes possèdent à la fois des motoneurones excitateurs libérant du glutamate et des motoneurones inhibiteurs qui libèrent du GABA (acide γ-aminobutyrique).
Figure 3 :Description de deux motoneurones dans le ganglion métathoracique d’un criquet, des principales branches dendritiques et des axones s’étendant dans le nerf latéral 3. Les corps cellulaires sont dans la partie externe du ganglion tandis que les dendrites sont dans la neuropile centrale (d’après Burrows, 1977) (Source : Structure and fonction – Chapman – Ed5-2013)
Les motoneurones inhibiteurs diminuent la force de contraction qui peut être produite par un muscle et augmentent son taux de relachement après la contraction. Ils peuvent agir directement sur les fibres musculaires mais peuvent également affecter de manière présynaptique l’activité des motoneurones excitateurs. Les motoneurones inhibiteurs innervent plusieurs muscles, souvent des paires antagonistes, et possèdent des axones particulièrement ramifiés. Par exemple, la patte d’un criquet est innervée par 70 motoneurones excitateurs et par seulement trois motoneurones inhibiteurs.
Les motoneurones excitateurs peuvent être subdivisés en 3 groupes : 1) lente ; 2) intermédiaire et 3) rapide en fonction de la vitesse de contraction musculaire qu’ils entraînent. Chez les insectes, la force et la rapidité de la contraction musculaire dépendent davantage du motoneurone individuel que de l’activité globale d’un grand nombre de neurones fonctionnellement homogènes, comme chez les vertébrés.
Interneurones
En général, comme leur nom l’indique, les interneurones jouent un rôle d’intermédiaire entre les neurones sensoriels et les neurones moteurs. La plupart des neurones du système nerveux central sont des interneurones. Des connexions synaptiques directes sont toutefois connues pour se produire entre les fibres afférentes (sensorielles) et les motoneurones. Bien qu’apportant une réponse rapide, ces voies monosynaptiques sont rares car cet arrangement entraîne une faible variabilité de la réponse.
Anatomiquement, les interneurones peuvent être divisés en deux groupes : les locaux et les intersegmentaires.
Les interneurones locaux sont limités à un seul ganglion, pouvant être stimulés ou non. On estime à 1 500 interneurones intraganglionnaires dans le ganglion métathoracique de la blatte Periplaneta, contre 200 interneurones interganglionnaires
Les interneurones segmentaires transmettent quant à eux des informations le long du cordon nerveux. Les fibres qui transportent des informations des ganglions antérieurs aux ganglions postérieurs sont appelées fibres descendantes, celles qui transmettent des informations au cerveau ou au ganglions antérieurs sont appelées fibres ascendantes. Ces neurones sont responsables de la coordination des activités des différents ganglions et provoquent ainsi la coordination de l’ensemble de l’insecte. Les axones de ces neurones peuvent se ramifier dans un ou plusieurs ganglions voisins ou non, ou transiter à travers des ganglions pour se ramifier uniquement dans des ganglions plus éloignés
Figure 4 : Innervation de muscles abdominaux de Teleogryllus par des neurones provenant de différents ganglions segmentaires – Seul les muscles d’un seul côté sont représentés (Source : Structure and fonction – Chapman – Ed5-2013)
L’un des interneurones intersegmentaires les plus étudiés est le détecteur de mouvement contralatéral (dont l’effet se manifeste du côté opposé au côté atteint) descendant (DCMD) du criquet. Ce neurone, s’étendant du cerveau au ganglion métathoracique, intervient dans le déclenchement de comportements de fuite en présence d’un danger (saut par exemple).
Le déclenchement d’un comportement de fuite fait intervenir un second interneurone entièrement situé dans le cerveau et connecté en partie avec le lobe optique (lobula) : le détecteur de mouvement géant lobula (LGMD). Le LGMD, particulièrement sensible aux stimuli visuels résultant de la détection d’objets mobiles sur la rétine des yeux composés (lien), envoie un signal nerveux au DCMD qui transmet à son tour le signal aux ganglions du thorax : prothoracique, mésothoracique et métathoracique. Ce signal est ensuite orienté vers un certain nombre de neurones excitateurs des muscles du tibia et du neurone inhibiteur commun de la musculature de la patte (figure 4).
Il fournit également une entrée inhibitrice au motoneurone du muscle adducteur coxal. L’activité dans le DCMD, telle que celle qui se produit lors de l’approche d’un objet en collision, est suffisante pour induire des manœuvres de décrochage et de plongée pendant le vol et peut jouer un rôle dans l’initiation d’autres comportement d’évasions et d’évitement des prédateurs.
Au sein d’un grand nombre de taxons comme les blattes, divers orthoptères, les larves de libellules et de drosophiles, voire chez tous les insectes, les cordons nerveux ventraux contiennent des axones appelés fibres géantes.
Chez la blatte par exemple, ces fibres géantes sont au nombre de 6 à 8 avec un diamètre de 20 à 60µm (en comparaison, les axones de la plupart des interneurones font moins de 5µm de diamètre). Le corps cellulaire de ces fibres est situé dans la ganglion abdominal terminal.
Chez la drosophile, les adultes ne possèdent quant eux que deux interneurones géants dont les corps cellulaires se situent dans le cerveau.
Certaines de ces fibres géantes s’étendent sur des longueurs considérables le long du cordon nerveux. Chez la blatte Periplaneta, les plus grosses fibres s’étendent du ganglion abdominal terminal au ganglion sous-œsophagien, soit sur la longueur du corps de l’insecte. Dans le ganglion abdominal terminal, chaque fibre géante se termine par une arborisation dendritique étendue au sein de laquelle des synapses sont formées avec des fibres nerveuses afférentes provenant du nerf cercal (qui innerve les cerques situés à l’extrémité de l’abdomen) (figures 5 et 6). D’autres fibres géantes se ramifient dans différentes parties du ganglion, suggérant qu’elles reçoivent des informations provenant de différents groupes de sensilles (organes sensoriels) (lien).
Figure 5 : Chez la blatte, les 3 ganglions thoraciques (méta-, méso- et prothoracique) contrôlent le mouvement des pattes. Des faisceaux d’axones assurent la communication entre les ganglions. Les neurones afférents des poils filiformes voyagent dans les nerfs cercaux jusqu’à une zone du ganglion (en gris) où ils se synapsent avec les interneurones (Source : Yager, 2010)
Les fibres géantes interviennent dans la transmission rapide d’informations sur de longues distances en raison de leurs taux de conduction élevés, de leur grande taille et de l’absence de synapses. A ce titre, elles jouent également un rôle important dans les mécanismes induisant les comportements de fuite.
Figure 6 : Intégration d’une fibre gante dans un ganglion abdominal terminal de blatte Periplaneta – a) Schéma montrant l’arbre dendritique et la position du corps cellulaire – b) Le neurone avec l’axone géant est directement connecté aux neurones sensoriels des cerques et des interneurones locaux (voies excitatrices et inhibitrices) (Source : Structure and fonction – Chapman – Ed5-2013)
Nerfs périphériques
De chaque ganglion rayonnent des nerfs périphériques constitués d’un grand nombre d’axones indépendants les uns des autres, ne se ramifiant pas et ne formant pas de synapses. Les axones de ces nerfs transmettent l’information provenant des neurones sensoriels périphériques (fibres afférentes) aux ganglions. Ceux distribuant les informations provenant du système nerveux central sont appelés fibres efférentes ou motrices.
Hormis les nerfs antennaires et cercaux constitués uniquement d’axones sensoriels, les nerfs périphériques contiennent à la fois des axones afférents et efférents diffusant symétriquement (homologie) dans un axe droite/gauche. Certains nerfs, comme le petit nerf médian, s’étend d’un ganglion thoracique à l’autre, se ramifiant transversalement aux stigmates et à certains muscles ventilatoires.
Neurone
Les neurones, ou cellules nerveuses, constituent l’unité fonctionnelle du système nerveux. Ce sont des cellules spécialisées pour transmettre des informations à d’autres cellules, nerveuses, musculaires ou encore glandulaires. Chaque cellule nerveuse est constituée d’un corps cellulaire contenant le noyau et le cytoplasme, de dendrites recevant les informations provenant d’autres neurones et d’un axone diffusant les messages nerveux aux autres cellules.
Il existe trois types de neurones : 1) les neurones monopolaires, constitués d’un seul axone dont une extrémité joue le rôle de dendrite ; 2) les neurones bipolaires, constitués d’un seul axone et d’une seule dendrite courte généralement non ramifiée recevant les stimuli provenant de l’environnement (organes sensoriels) ; 3) les neurones multipolaires, constitués d’un seul axone et d’une multitudes de dendrites, se rencontrent principalement dans les ganglions du système nerveux sous forme d’interneurones ou de neurones moteurs) (figure 7).
Quand un neurone reçoit ou émet un message, il transmet des impulsions électriques le long de l’axone (pouvant mesurer plus d’un mètre). Le message électrique, ou influx nerveux, repose sur l’ouverture et la fermeture de canaux ioniques qui induisent le transfert d’ions à travers la membrane de la cellule, produisant ainsi un courant électrique.
Figure 7 : représentation des différents types de neurones (les flèches indiquant la direction de l’information nerveuse) (Source : Structure and fonction – Chapman – Ed5-2013)
The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
Intestin postérieur (proctodeum) également d’origine ectodermique
Contrairement au tube digestif des vertébrés couvert d’une paroi ayant de nombreuses couches (muqueuse, sous-muqueuse, musculeuse et tunique externe), celui des insectes est constitué d’une couche unique de cellules épithéliales que délimite une membrane basale et des muscles striés.
Il forme un conduit linéaire continu de la bouche à l’anus. Des régionalisations et des zones de spécialisation sont présentes : diverticules au niveau de l’intestin moyen, caecum au niveau de l’intestin moyen, tubes de Malpighi à la jonction entre les intestins moyen et postérieur.
Chez les insectes se nourrissant de matières peu solides, voire de fluides, comme les Hémiptères (suceurs de sèves), les larves de Neuroptères (fourmilions par exemple) qui avalent une proie liquéfiée par digestion externe (lien) ou encore les larves d’Hyménoptères sociaux dont les adultes prémâchent la nourriture, un rétrécissement sépare l’intestin médian à l’intestin postérieur.
Figure 1 : Description des différentes subdivisions du tube digestif des insectes (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
Diversité anatomique des tubes digestifs chez les insectes : figure 11 en bas de page.
Origine embryonnaire
D’origine ectodermique, les parois de l’intestin antérieur et postérieur sont couvertes par la cuticule qui, sécrétée par des cellules épidermiques, constitue la continuité avec l’exosquelette de l’insecte. Ces parois sont évacuées puis renouvelées avec l’ensemble de l’exosquelette lors de chaque mue.
L’intestin moyen n’est quant à lui pas recouvert de cuticule, mais de nombreux insectes sécrètent toutefois une membrane uni-couche non-cellulaire (enveloppe péritrophique) dont le rôle est d’isoler les cellules épithéliales de la nourriture.
Intestin antérieur
Les cellules de l’intestin antérieur sont généralement aplaties et indifférenciées du fait qu’elles n’interviennent ni dans l’absorption des nutriments ni dans la sécrétion d’enzymes. La membrane cuticulaire est en générale peu sclérifiée mais uniquement constituée d’une endocuticule et d’une épicuticule (lien). Chez de nombreux insectes, la surface de l’intestin antérieur est parfois recouverte d’épines ou de dents sclérifiées. De forme et de disposition variables selon les espèces, ces excroissances pointent vers l’arrière afin de faciliter le transit de la nourriture (figure 2).
Figure 2 : Adaptations de l’intestin antérieur pour la digestion mécanique des aliments – (a) Coupe sagittale à travers l’intestin antérieur d’un criquet montrant le motif d’épines cuticulaires. Les agrandissements montrent les détails des épines. Dans le proventricule, les épines sont remplacées par des plaques sclérotisées plus grandes avec des dents dirigées vers l’arrière sur les bords postérieurs (d’après Williams, 1954). (b) Coupe longitudinale de l’intestin antérieur de blatte Periplaneta montrant le développement du proventricule pour former
un appareil de broyage (d’après Snodgrass, 1935) (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
L’intestin antérieur est subdivisé en quatre parties : pharynx, œsophage, jabot, proventricule.
Le pharynx intervient dans l’ingestion de la nourriture une fois celle-ci amenée par la bouche : il est constitué d’une musculature développée permettant la progression de celle-ci vers l’œsophage
L’œsophage relie le pharynx au jabot. Souvent mal caractérisé chez les insectes hémimétaboles (métamorphose incomplète – lien), mais il forme un long tube élancé traversant le thorax et les muscles du vol chez les insectes holométaboles (métamorphose complète –lien). Les espèces se nourrissant de plantes résineuses possèdent souvent une adaptation singulière de l’œsophage : des diverticules simples ou appariées y jouent un rôle dans le stockage de la résine (exemple : chenilles du genre Myrascia qui se nourrissent de Myrtacées (figure 3) ou larves de tenthrèdes du genre Neodiprion qui se nourrissent de pins et dont les diverticules sont dotés de puissants muscles circulaires leur permettant d’éjecter le contenu du jabot en réponse aux attaques de prédateurs)
Le jabot est un organe de stockage, extensible chez la plupart des insectes (figure 3). Chez les Lépidoptères et les Diptères, il forme un diverticule latéral de l’œsophage. Plissées longitudinalement et transversalement, ses parois se distendent au fur-et-à-mesure que le jabot se remplit, d’où une importante augmentation de son volume. Cas particulier, chez les blattes Periplaneta, le jabot, bien que vide, garde un volume constant en se remplissant d’air. La capacité du jabot à contenir du liquide réside dans l’imperméabilité de la muqueuse cuticulaire aux molécules hydrophiles (molécules ayant une affinité avec l’eau vs molécules hydrophobes)
Le proventricule présente une grande diversité de forme. Il s’ouvre par une simple valve à l’extrémité antérieure de l’intestin moyen, pénétrant parfois sur une courte distance dans la lumière de ce segment. Chez les criquets (Acrididae – Orthoptères), le proventricule forme une constriction limitant le passage des aliments solides vers l’intestin moyen tout en laissant diffuser les liquides dans les deux sens. Les fourmis possèdent un proventricule spécialisé séparant la nourriture partiellement digérée de la nourriture stockée dans le jabot. Ce mécanisme intervient lors de la trophallaxie (échange de nourriture entre deux individus par régurgitation). Le proventricule est également spécialisé chez les abeilles mellifères (qui fabriquent du miel) (figure 4) ce qui leur permet de retenir le nectar dans le jabot tout en laissant passer les grains de pollen vers l’intestin moyen. Chez les Orthoptères, les blattes (Blattodea) et certains Coléoptères, les proventricules sont constitués de plaques cuticulaires ou de dents intervenant dans le broyage d’aliments solides (figure 2).
Figure 3 : Types de stockage de la nourriture dans l’intestin – zones hachurées – (a) à (d) Stockage dans l’intestin antérieur – (a) Une chenille typique – (d) une chenille qui se nourrit de plantes résineuses et stocke la résine dans le diverticule œsophagien. (e) – (f) Stockage dans l’intestin postérieur – (g) Stockage dans l’intestin moyen – Figure 4 : Intestin antérieur d’une abeille ouvrière en coupe longitudinale montrant le développement du proventricule. La partie dirigée vers l’avant permet à l’insecte d’extraire les grains de pollen du nectar de la culture – la partie postérieure faisant saillie dans l’intestin moyen formant une valve (d’après Snodgrass, 1956) (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
Intestin moyen
La partie tubulaire de l’intestin moyen (également appelée ventricule) présente de nombreuses différenciations régionales marquées à la fois au niveau anatomique (Hétéroptères par exemple) et cellulaire (Diptères par exemple). Chez une multitude d’insectes, l’intestin moyen comporte des caecums dans la partie antérieure (figure 1 ci-dessus). Le nombre de caeca varie selon les taxons : 2 chez les grillons (Gryllidae) et certaines larves de Diptères, 6 chez les criquets (Acrididae), 8 chez les blattes (Blattodea) et les larves de Culicidae (moustiques – Diptères).
Les cellules de l’intestin moyen (entérocytes) sont impliquées dans la production et la sécrétion d’enzymes digestives et dans l’absorption des nutriments. La majorité d’entre elles, rectangulaires, forment des « colonnes » dont la membrane luminale (en contact avec la lumière de l’intestin) porte des microvillosités augmentant ainsi la surface d’échange parfois d’un facteur 100 (figure 3.5).
Figure 5 : Épithélium de l’intestin moyen d’une larve de Drosophila melanogaster et pH à l’intérieur de l’intestin. Le contenu du segment antérieur et de la partie antérieure du segment postérieur se situe entre une alcalinité neutre à légère (pH> 7 et <8) – le segment médian est très acide (pH <3,0) – et la partie postérieure du segment postérieur est fortement alcaline (pH> 10) – HG : intestin postérieur; MT : tubes de Malpighi – (b) Représentation schématique d’une coupe transversale de la région postérieure de l’intestin moyen postérieur larvaire montrant la disposition de la membrane péritrophique, des cellules épithéliales de la membrane basale, des fibres musculaires, des cellules régénératives ou des cellules souches (marquées S) – (c) Organisation générale de deux cellules épithéliales adjacentes. La flèche indique de très petits espaces intercellulaires latéraux – BEL : labyrinthe extracellulaire basal – BM : membrane basale – CM : muscle circulaire – JC : complexe jonctionnel – LM : muscle longitudinal – M : mitochondries – MV : microvillosités – PM : membrane péritrophique – SB : barrière séreuse – T : trachéole (reproduit de Shanbhag et Tripathi, 2009) (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
Les autres types de cellules prédominants dans l’épithélium de l’intestin moyen sont les cellules dites entéroendocrines sécrétant des hormones de régulation et les cellules souches intestinales à l’origine des cellules différenciées de ce segment (figure 6). Les cellules souches forment souvent des amas appelés « nidus ». Chez certains Coléoptères, les nidus sont limités aux cryptes des cellules souches (cellules invaginées formant des cryptes) sous forme de petites papilles (figure 6).
L’intestin moyen des larves de Lépidoptères possède d’autres types de cellules appelées « cellules en coupe » dont la membrane luminale, présentant des microvillosités, est invaginée pour créer une cavité en forme de ballon (figure 7). Ces cellules assurent l’alcalisation de ce segment (pH basique – ph<7). Une large cavité médiane est présente chez les éphémères (Éphéméroptères), les Plécoptères et les Trichoptères sans que sa fonction soit connue.
Chez tous les insectes, les cellules souches des larves prolifèrent et se différencient en entérocytes et en cellules entéroendocrines durant la métamorphose pour former l’intestin moyen de l’adulte. Les cellules souches continueront à jouer ce rôle durant toute la vie de l’insecte.
L’épithélium de l’intestin moyen de l’adulte a un temps de renouvellement de 1 à 2 semaines, les anciennes cellules étant expulsées dans la lumière intestinale. Ce phénomène est favorisé par la présence de bactéries symbiotes stimulant la production d’oxygène par les entérocytes, et augmentant la prolifération et la différenciation des cellules souches.
La couche épithéliale larvaire est cependant entièrement remplacée lors de la métamorphose par un épithélium nymphal (adulte) qui persiste tout au long de la vie adulte de l’insecte.
Chez les larves de drosophile, (Drosophila melanogaster – Diptères) la croissance de l’intestin moyen résulte principalement de l’augmentation de la taille des cellules le constituant.
Chez les larves de Lépidoptères, un pic de division de cellules souches a lieu avant chaque mue et les cellules nouvellement différenciées s’intercalent entre les cellules de l’épithélium préexistant. Durant la métamorphose, l’épithélium de l’intestin moyen des chenilles subit une apoptose (mort cellulaire programmée – autodestruction) et une autophagie (recyclage des éléments cellulaires) puis les déchets métaboliques sont rejetés après l’émergence du papillon adulte.
Figure 6 : (a) Groupe de cellules souches (nidus) à la base de l’épithélium de l’intestin moyen montrant la différenciation des entérocytes (d’après Fain-Maurel et al., 1973). (b) Crypte de l’intestin moyen d’un coléoptère s’étendant à travers la couche musculaire pour former une papille (d’après Snodgrass, 1935). Figure 7 : (a) Cellule à gobelet (d’après Cioffi, 1979) (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
Enveloppe péritrophique
L’intestin moyen de la plupart des insectes est séparé de la lumière par une structure non cellulaire appelée enveloppe péritrophique (membrane péritrophique ou matrice péritrophique). La composition et le mode de formation de cette enveloppe varient selon les insectes, mais elle comprend généralement un gel de protéoglycane scouvrant une structure rigide de microfibres de chitine (un des constituants de la cuticule) (figure 8 ci-dessous).
L’enveloppe péritrophique est dite de type I si elle est produite sur toute la longueur de l’intestin moyen, ou de type II si elle n’est produite que dans la partie antérieure de l’intestin moyen. De nombreuses variations existent au sein de chacun de ces deux types.
Le type I se rencontre chez les Coléoptères, les Blattodea, les Éphéméroptères, les Hyménoptères, les Odonates, les Orthoptères, les Phasmides, les larves de Lépidoptères ou encore les Diptères adultes.
Chez un criquet qui se nourrit activement, une nouvelle enveloppe est produite toutes les 15 minutes de sorte que les enveloppes successives se superposent. Les couches les plus anciennes sont éliminées avec la nourriture au fur-et-à-mesure que de nouvelles sont produites. La résistance mécanique de l’enveloppe péritrophique varie en fonction des espèces. Par exemple, les criquets (Acrididae) granivores du genre Locusta synthétisent une enveloppe fine et délicate tandis que l’enveloppe des espèces polyphages comme celles du genre Schistocerca, est plus épaisse et dure.
Type II : l’enveloppe péritrophique de type II est caractéristique des Dermaptères (perce-oreilles – lien), des Isoptères (termites – lien), de certains Lépidoptères et des larves de Diptères. Chez les larves de moustiques Aedes, les 8 à 10 anneaux de cellules les plus antérieurs de l’épithélium de l’intestin moyen (seulement 300 à 400 cellules) produisent toute l’enveloppe péritrophique constituée d’une seule couche de 0,7mm d’épaisseur. Chez Forficula (Dermaptères), l’enveloppe est constituée de plusieurs couches.
L’enveloppe péritrophique s’apparente à un tamis moléculaire dans lequel les pores sont des canaux aqueux traversant le gel de protéoglycane. La microscopie électronique permet de déterminer les dimensions de ces spores et de constater leur perméabilité à des particules de 21 à 36nm, telles des ions inorganiques, de petites molécules organiques (sucres, acides aminés, etc.), de peptides, voire de petites protéines. Cette enveloppe constitue en revanche une barrière efficace au passage des lipides, des polysaccharides (polymères de glucides) et d’autres grosses molécules.
Elle protège également la paroi intestinale des dommages causés par les matières ingérées comme des particules alimentaires abrasives, des agents pathogènes (résistance de jeunes larves d’abeille âgées de 3 jours à la bactérie Paenibacillus en partie retenue par la membre péritrophique) ou encore de certaines toxines (tanins de plantes par exemple). Cependant, bien que certaines molécules de tanins soient plus petites que les pores, le mécanisme permettant à la membrane de les retenir n’est pas encore élucidé. Chez Locusta, la membrane épitrophique n’est pas perméable à l’acide tannique. Lors de l’ingestion de trop d’aliments contenant cette substance, les insectes risquent des lésions épithéliales de l’intestin moyen pouvant entrainer leur mort.
Enfin, l’enveloppe joue un rôle dans la compartimentation des activités enzymatiques et l’entretien d’un écoulement à contre-courant de fluides augmentant l’efficacité de leur absorption.
Le rôle de l’enveloppe péritrophique dans la protection mécanique est démontré lors d’études sur des larves mutantes de Bombyx mori incapables de la produire.
L’enveloppe péritrophique est absente chez certains insectes comme les moustiques adultes, n’apparaissant qu’à la suite d’un repas chez les femelles, les papillons se nourrissant de nectar, et les Hémiptères et les Thysanoptères (trips) où une couche de lipoprotéines (membrane périmicrovillaire) la remplace.
Figure 8 : Description anatomique de l’enveloppe péritrophique dans l’intestin moyen (Source : http://entnemdept.ufl.edu – modifié par B. Gilles)
Intestin postérieur
L’intestin postérieur est généralement différencié en trois régions : pylore, iléon et rectum (figure 1 ci-dessus).
Comme pour l’intestin antérieur, il est recouvert d’une couche de cuticule, toutefois plus fine et plus perméable que celle de l’intestin antérieur.
Le pylore constitue la zone où se mélangent les résidus alimentaires de l’intestin moyen et les sécrétions des tubules de Malpighi qui y débouchent. Chez de nombreux insectes, il est délimité par un muscle circulaire et agit donc comme un sphincter contrôlant le passage de la nourriture de l’intestin moyen vers l’intestin postérieur et prévenant le reflux du contenu de l’intestin postérieur vers l’intestin moyen.
L’iléon est un tube étroit situé entre le pylore et le rectum. La partie distale de l’iléon pouvant parfois être différente, cette partie est alors appelée côlon. L’iléon peut également être élargi dans sa partie postérieure pour abriter des microorganismes symbiotes formant la « panse » chez les termites et la « chambre de fermentation » chez certains Scarabaeidés (bousiers par exemple) (figure 3 ci-dessus). La cuticule tapissant ces chambres forme des épines allongées ou des lobes bulbeux servant probablement à la fixation des microorganismes.
Le rectum consiste en un sac élargi protégé par un épithélium mince à l’exception de certaines régions, plus épaisses, comme les coussinets rectaux composés des cellules épithéliales en forme de colonne et où a lieu l’absorption de l’eau.
Le rectum est également un site d’échanges des gaz respiratoires chez certains insectes aquatiques comme les larves d’Odonates chez lesquelles la partie postérieure du rectum forme une chambre branchiale constituée de filaments ramifiés (figure 9).
Figure 8 : Branchies trachéales dans le rectum d’une larve de libellule (d’après Tillyard, 1917; Mill et Pickard, 1972). (a) Coupe longitudinale à travers l’abdomen – les nombres indiquant les segments abdominaux – (b) Coupe transversale à travers la chambre branchiale – (c) Coupe transversale à travers le segment abdominal 6 montrant le muscle sous-intestinal – (d) Détail de l’épithélium des branchies montrant les trachées (d’après Schmitz et Komnick, 1976) (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
Musculature du tube digestif
Les muscles du tube digestif se divisent en deux catégories : les muscles viscéraux extrinsèques qui sont attachés à la paroi corporelle et sont insérés dans l’intestin, et les muscles viscéraux intrinsèques qui ne sont associés qu’à l’intestin.
Les muscles viscéraux intrinsèques comprennent des muscles striés circulaires et longitudinaux s’étendant le long du tube digestif. Ils sont bien développés autour du pharynx et du proventricule, a contrario, moins développés le long de l’intestin moyen.
Les muscles viscéraux extrinsèques ressemblent quant à eux aux muscles squelettiques. Localisés dans l’intestin antérieur et postérieur, ils interviennent dans la dilatation de l’intestin. Dans la partie antérieure, ils jouent un rôle de pompe aspirant les fluides vers l’œsophage. Dans l’intestin postérieur, ils refoulent l’eau vers les chambres branchiales des larves de libellules par exemple. (figure 8).
Passage de l’aliment à travers le tube digestif
L’aliment est déplacé à l’aide des muscles de la pompe pharyngée puis véhiculé le long de l’intestin par péristaltisme (ensemble des contractions musculaires permettant la progression d’un contenu à l’intérieur d’un organe creux).
Contrôlés à partir du système nerveux antérieur, ces mouvements sont modulés par diverses neurohormones. Par exemple, lorsqu’un criquet Locusta (Acrididés) se nourrit après une période de jeûne de plusieurs heures, la nourriture solide reste dans l’intestin antérieur jusqu’à ce que celui-ci soit rempli et distendu. Le bol alimentaire débute alors son déplacement vers l’arrière. Chez les femelles de mouches hématophages et de celles se nourrissant de nectar, la destination de l’aliment est déterminée par sa composition chimique détectée par les récepteurs des pièces buccales : la stimulation avec des sucres à des concentrations élevées dirige le repas vers le jabot (figure 3) tandis qu’une stimulation avec des protéines entraîne le repas vers l’intestin moyen.
Dans l’intestin moyen de la plupart des insectes, le déplacement de la nourriture est facilité par l’enveloppe péritrophique. Les insectes hématophages comme les punaise Rhodnius et Cimex, les moustiques ou les taons (Tabanidae) qui prennent des repas peu fréquents stockent la nourriture dans l’intestin moyen antérieur (figure 3), l’eau est absorbée directement.
Les mouvements de l’intestin postérieur sont importants dans l’élimination du matériel non digéré. Chez le criquet Schistocerca (Acrididae), l’iléon forme généralement un coude en « S » provoquant le déchirement de la membrane péritrophique qui, mélangée avec le bol alimentaire, forme ainsi un culot fécal. Les contractions de la partie distale de l’iléon et du rectum forcent ce culot à sortir par l’anus. Chez les sauterelles, les excréments sont enfermés dans l’enveloppe péritrophique alors que chez les chenilles, l’enveloppe est déchirée dans l’intestin postérieur.
Le temps nécessaire pour que la nourriture traverse l’intestin est variable. Chez une sauterelle avec un accès continu à la nourriture, les liquides atteignent l’intestin moyen et sont absorbés dans les 5 minutes suivant le début du repas, les particules solides mettront 15 minutes ou plus. La nourriture quitte l’intestin avant 90 minutes environ, repoussée par le repas suivant (figure 10). En l’absence de second repas, l’intestin antérieur est complétement vide 5 heures après l’ingestion, et l’intestin moyen après 8 heures environ.
Chez la blatte Periplaneta (Blattodea), les aliments solides traversent l’intestin en 20 heures environ, et, en cas de jeûne, de l’aliment peut y être retrouvé après plusieurs jours. Chez les termites qui se nourrissent de bois et de matières végétales, des fragments de nourriture sont conservés dans l’intestin postérieur durant de longues périodes afin que les microorganismes symbiotiques aient le temps de dégrader la cellulose.
Figure 10 : Déplacement de la nourriture dans l’intestin d’une sauterelle avec accès continu à la nourriture (d’après Baines et al., 1973) (Source : Insect – Structure and function – Chapman Ed5 – modifié par B. Gilles)
Diversité anatomique des tubes digestifs chez les insectes
Figure 11 : A) Structure intestinale généralisée des insectes. L’intestin antérieur et l’intestin postérieur sont bordés d’une couche de cuticule (ligne noire épaisse), l’intestin moyen sécrète une matrice péritrophique (ligne pointillée) – (B) à (M) Structures intestinales d’insectes d’ordres différents (Source : Engel & Moran, 2013 – Modifié par B. Gilles)
Bibliographie
The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
Engel P. & Moran N. (2013) : The gut microbiota of insects – Diversity in structure and function. FEMS Microbioloy Reviews, 37(5), 699-735 (lien)
Recommandation d’ouvrages sur cette thématique
The Insects : Structure & Function (5th edition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
Evolution of the Insects(David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
Avec près de 100 000 espèces décrites, le règne des champignons constitue l’un des groupes les plus diversifiés des eucaryotes (organismes cellulaires à noyaux) et ayant conquis l’ensemble des écosystèmes terrestres et aquatiques. Ce nombre représente une infime fraction de la diversité connue estimée, quant à elle, entre 1,5 millions et 5 millions d’espèces.
Une des propriétés des champignons est leur propension à former des interactions intimes avec d’autres groupes d’organismes tant animaux que végétaux : 21% des espèces sont associées à des algues (lichens) et 8% entretiennent des relations symbiotiques avec des végétaux (mycorhizes).
Les organismes n’entretenant aucune association fongique sont rares, voire inexistant. Les champignons sont de ce fait des acteurs essentiels dans le maintien des équilibres des écosystèmes.
Les insectes, avec plus de 1,1 millions d’espèces décrites, représentent, quant à eux, le groupe d’organismes eucaryotes le plus diversifié. Ils sont connus pour entretenir des relations avec de nombreux taxons de champignons notamment de type 1) endosymbiotiques mutualistes intervenant dans la nutrition (fourmis champignonnistes par exemple) ; 2) commensales ; 3) pathogènes avec des effets prononcés sur les populations.
L’étude des relations entre champignons entomopathogènes et les insectes reste l’une thématique de l’entomologie les moins bien étudiées et offrant probablement l’un des plus grands réservoirs d’espèces fongiques à découvrir.
La première difficulté que rencontre les champignons entomopathogènes est de pénétrer le corps du futur insecte hôte recouvert d’un exosquelette composé en grande partie de chitine. Une fois dans l’organisme, le type développement des champignons dépend du groupe auquel il appartient : formation d’hyphes (succession d’articles composant le mycélium) semblables à celles des levures et multiplication par bourgeonnement (Metarhizium et Beauveria – Hypocrales – Ascomycota), production de protoplastes (fusion des cellules par perte de la membrane externe) (Entomophthoromycota) ou de des filaments non cloisonnés (forme d’hyphes) (Oomycota, Chytridiomycota ou des espèces du genre Entomophthora).
La majorité des champignons entomopathogènes tuent leur hôte avant l’émission des spores (équivalent de la graine chez les plantes) : Hémibiotrophes. D’autres espèces sporulent quant à elle alors que l’hôte demeure en vie (phylum des Entomophthoromycota) : biotrophes.
Dans le milieu aquatique, la transmission à l’hôte est effectuée de manière active, les spores (zoospores) sexuées possèdent un flagelle les rendant mobiles (Chytriodiomycota et les Oomycota par exemple). Les spores peuvent être également de type mitotique asexué, relâchées passivement dans l’environnement, ils sont appelés conidies quel que soit le taxon.
Les spores sont nommées en fonction du groupe auquel ils appartiennent : zygospores (Zygomycètes), basidiospores (Basidiomycètes) et ascospores (Ascomycètes) (figure 1 ci-dessus).
La morphologie et le comportement de germination des spores interviennent dans la systématique et la classification des différents groupes de champignons.
Principaux groupes des champignons entomopathogènes
Oomycètes
Microsporidies
Chytridiomycètes
Zygomycètes
Basidiomycètes
Ascomycètes
1. Oomycètes
1.1. Généralités
Considérées parmi les champignons pour leurs similitudes écologiques et morphologiques avec les champignons, des études phylogénétiques (2006) ont confirmées que les Oomycètes étaient associés au groupe des Stramenophila, un groupement d’organismes morphologiquement divers comme les Hyphochtriomycota et les Labyrinthulomycota.
Photo 1 : larve d’insecte aquatique mort après infestation d’un champignon Oomycète Saprolenia sp. (Source : Encyclopedia Britannica)
Les Oomycètes possèdent des caractères biologiques spécifiques comme des spores à paroi épaisse (oospores) possédant deux flagelles (biflagélé), un flagelle en guirlande dirigée vers l’avant et un second plus court dirigé vers l’arrière (figure 1B). Au niveau cellulaire, ils présentent des mitochondries à crêtes tubulaires contrairement à celles des champignons qui présentent des crêtes aplaties en forme de plaque. De plus, leurs parois cellulaires contiennent de la cellulose contre de la chitine chez les champignons (matériaux constituant également l’exosquelette des insectes).
Les Oomycètes sont des parasites et des saprophytes (se nourrissant de tissus morts et de matières organiques). En tant qu’agents pathogènes, les Oomycètes sont capables d’infecter un large éventail d’hôtes tels que les algues, les plantes, les protistes, les champignons, les arthropodes (photo 1 ci-contre) et les vertébrés y compris les humains. Certains genres sont des agents phytopathogènes bien connus comme les membres du genre Phyphthora : agent responsable de la famine irlandaise de la pomme de terre, également à l’origine de la mort subite du chêne qui affecte actuellement des millions d’arbres. Les Oomycètes infectent aussi des arthropodes marins comme le homard et les crevettes.
1.2. Espèces entomopathogènes
Les Oomycètes entomopathogènes sont au nombre de 12 espèces réparties en six genres : Lagenidium (L. giganteum), Leptolegnia (L. caudata et L. chapmanii), Pythium (P. carolinianum, P. sierrensis et P. flevoense), Crypticola (C. clavulifera, C. entomophaga), Couchia (C. amphora, C. linnophila et C. circumplexa) et Aphanomyces (A. laevis).
Les Oomycètes ont été identifiées uniquement chez des larves aquatiques de Diptères des genres suivants : Aedes, Anopheles, Chironomus, Culex, Forcipomyia, Glyptotendipes, Mansonia, Ochlerotatus, Pentaneura, Polypedilum, Tendipeset Uranotaenia.
Un seul un cas d’infections chez une espèce autre qu’un Diptère a été décrit chez Crypticola entomophaga (Thrichoptera) dont les larves sont également aquatiques.
Figure 1 : Cycle de développement de Lagenidium giganteum (Oomycètes) A) La zoospore adhère et pénètre dans la cuticule, déclenchant l’infection – B) le mycélium commence à se développer et à proliférer dans le corps de la larve – C) zoosporangium libérant des zoospores asexuées – D) oospores (partie sexuelle du cycle) – E) oospore se détache des hyphes et s’installe dans l’environnement – F) libération des zoospores sexuelles (Source : Araujo & Hughes, 2016)
L’espèce d’Oomycètes la plus connue et étudiée est L. giganteum, un parasite facultatif des larves de moustiques (figure 1 ci-contre). Des études ont décrit que le comportement de nage des zoospores vers la surface de l’eau était davantage associé à la forme cellulaire et au centre de gravité qu’à une réponse sensorielle.
D’autres genres comme Leptolegnia, Pythium, Crypicola et Aphanomyces ont cependant peu fait l’objet d’intérêts scientifiques.
Les espèces Leptolegniacaudata et L. chapmanii ont par exemple été isolées de larves de moustique Aedes aegypti, vecteur de la fièvre jaune dont le cycle de vie dont le cycle de vie a été particulièrement bien décrit. Des tests d’infestations chez des Odonata, des Trichoptera, des Coleoptera, des Plecoptera et des Cladocera ont été entrepris sans succès suggérant une spécificité du pathogènes envers les Diptères.
Les espèces du genre Pythium se retrouvent principalement sous forme d’organismes vivant dans le sol ou d’agents pathogènes des plantes. Seules trois espèces sont connues pour infecter des larves d’insectes.
Concernant Aphanomyces, bien que quelques cas d’épizooties ont été signalées, peu d’études ont été publiées sur ce genre.
2. Microsporidies
2.1. Généralités
Traditionnellement, les Microsporidies sont classées dans le phylum des « parasites sporozoaires » (parasites d’organismes pluricellulaires). U nombre croissant d’études soutiennent l’hypothèse selon laquelle les Microsporidies seraient plutôt des champignons parasites endocellulaires.
La caractéristique la plus remarquable de ce groupe est la taille des spores compris entre 1 et 40µm agissant comme une seringue injectant son matériel protoplasmique dans l’hôte au niveau de l’apex à la manière d’un « retournement du doigt d’un gant ».
La gamme d’hôtes de la plupart des espèces de Microsporidies est relativement restreinte : animaux domestiques, poissons, amphibiens, reptiles, oiseaux et mammifères.
2.2. Espèces entomopathogènes
Les Microsporidies sont un groupe d’agents pathogènes comprenant 143 genres avec plus de 1 200 espèces, dont 69 genres infectant, à ce jour, 12 ordres d’insectes. La majorité (42 genres) infectent des Diptères, 5 genres des Ephéméroptères et des Coléoptères, 4 genres des Lépidoptères, 3 genres des Trichoptères, les Orthoptères, les Odonates et les Siphonaptères étant infestés par 2 genres, les Hyménoptères, les Thysanoures et les Isoptères par un seul genre.
Le groupe le plus commun est celui du genre Amblyospora pouvant infecter 79 espèces de mouches réparties en 8 genres. Le cycle de vie de Amblyospora est complexe nécessitant un hôte copépode intermédiaire et deux générations de moustiques afin de réaliser entièrement son cycle.
Un autre groupe important parmi les Microsporidies est le genre Nosema (figure 2 ci-dessous). Certains auteurs le considèrent comme le plus largement distribué et étant responsable de la majorité des infections microsporidiennes chez les espèces de Lépidoptères. En infectant les abeilles, les espèces N. bombycis et N. ceranae sont à l’origine d’impacts écologiques et économiques majeurs. La contamination se fait par l’ingestion de spores par des abeilles adultes qui vont ensuite rejoindre puis pénétrer les cellules épithéliales de l’intestin moyen des hôtes. La libération dans l’environnement de nouvelles spores infectieuses se fait avec les excréments, spores qui seront à leur tour ingérées par d’autres abeilles. La structure de la paroi cellulaire des spores est composée de 3 couches leur permettant de résister aux conditions de l’environnement.
Figure 2 : Cycle de développement de Nosemia sp. A) ingestion des spores – B) spores atteignent l’intestion de l’abeille où ils s’activent en injectant des sporoplasme dans les cellules hôtes – C) multiplication cellulaire – D et E) transition du sporoplasme au spore – F) les spores sont à nouveau libérées dans l’intestion et se répandent dans les excréments de l’abeille ou réinfecteront le même hôte (Source : Araujo & Hughes, 2016)
3. Chytridiomycètes
3.1 Généralités
Le groupe des Chytridiomycètes est considéré comme la lignée divergente la plus ancienne avec les champignons (Dévonien inférieur : 400 millions d’années). Chytridiomycota est le seul taxon parmi les champignons à posséder des cellules mobiles durant son cycle de vie. Ces zoospores mobiles et aquatiques sont équipées d’un seul flagelle dirigé vers l’arrière : évolution convergente avec les Oomycètes.
La majorité des Chytridiomycètes sont des organismes saprophytes notamment dans les eaux douces et les sols humides, mais aussi de certaines espèces marines. Un nombre important d’espèces sont cependant connues pour parasiter des plantes, des animaux, des rotifères, des tardigrades, des protistes ainsi que d’autres champignons. Les cas de parasitisme chez les insectes sont particulièrement rares.
3.2 Espèces entomopathogènes
Les Chytridiomycètes regroupent 4 genres entomopathogènes : Myiophagus (M. ucrainicus), Coelomycidium (C. simulii), Myrmicinosporidium (M. durum) et le genre le plus diversifié Coelommoyces. Les cas d’infections ont été principalement été recensés chez les Diptères.
Figure 3 : Cycle de développement de Coelomyces phosphorae (Chytridiomycètes) A) les zoospores s’attachent et pénètrent dans la cuticule du copépode – B) développement et dispersion des gamètes dans l’environnement – C) fusion des gamètes compatibles à l’intérieur du copépode ou dans l’environnement – D) formation de zygote et fixation à la cuticule de la larve de moustique – E) colonisation, développement et formation de sporanges – F) libération de sporanges dans l’environnement après la mort de la larve – G) méiose et libération de zoospores asexuées – H) migration vers les ovaires si la larve reste vivante, le moustique femelle pondra alors des sporanges fongiques au lieu d’œufs (Source : Araujo & Hughes, 2016)
Le genre Myiophagus a été décrit chez des larves de Diptères et des cochenilles. En 1978, Doberski & Tribe ont signalé l’espèce Catenaria auxiliaris chez des larves de Coléoptères, cependant, les auteurs ne sachant pas si le champignon a colonisé l’insecte après sa mort (saprophytisme) ou si celui-ci a infecté l’insecte de son vivant, C. auxiliaris n’est actuellement pas considéré comme un entomopathogène.
Le genre Coelomycidium est connu pour infecter spécifiquement les mouches de la famille des Simuliidae. Cette pathologie est caractérisée par des larves remplies de sporanges sphériques dans toute la cavité corporelle.
Les espèces de Coelomomyces sont particulièrement bien connues car leurs hôtes sont d’importants vecteurs de maladies humaines (moustiques Anopheles, Culex et Aedes). Les espèces de ce genre peuvent contaminer les œufs, les larves et les adultes.
Chez certaines espèces comme C. phosphorae, un copépode est nécessaire pour achever l’ensemble du cycle de vie (figure 3 ci-contre).
Dans certains cas, le champignon ne tue pas les larves de moustiques, au contraire, il demeure à l’intérieur de l’insecte jusqu’au stade adulte où il colonise les ovaires des femelles. Après le premier repas sanguin de la femelle, les hyphes mûrissent pour devenir des sporanges à l’origine de la production de zoospores. Ainsi, au lieu de pondre des œufs, la femelle « pond » des sporanges remplis de zoospores prêts à infecter de nouvelles larves.
4. Zygomycètes
4.1. Généralités
Le groupe des Zygomycètes est traditionnellement subdivisé en deux classes : les Zygomycètes et les Trichomycètes partageant des caractéristiques communes comme le mycélium cénocytaire (sans septation régulière), la reproduction asexuée et l’absence de cellules à flagelles et de centrioles.
Leur principale caractéristique générale est la production d’une spore de repos à paroi épaisse (zygospore) dans un zygosporange formé après la fusion de deux hyphes spécialisés appelés gamétanges.
Le groupe est écologiquement diversifié et largement distribué, sous forme de saprotrophes dans le sol et les excréments (Kickxellomycotina, Mucoromycotina et Zoopagomycotina), dans le pain, les fruits et les légumes pour certains.
Photo 2 : Sporodiniella umbellata développé sur un insecte (Source : twitter.com/popos07)
La famille des Zoopagomycotina, également appelée « champignons piégeurs de nématodes », regroupe les espèces mycoparasitaires les plus connues. Celle des Mucoromycotina est quant à elle la plus diverse au sein des Zygomycètes. Sporodiniella umbellata est la seule espèce se rencontrant sur divers insectes comme les Membracidae (Hémiptères s’alimentant de la sève des plantes) (Photo 2).
Les Trichomycètes se rencontrent exclusivement dans les tubes digestifs de divers arthropodes. Cependant, leur présence n’inflige aucun dommage à leur hôte et la nature de leur relation n’est pas entièrement comprise.
L’un des groupes les plus importants de tous les entomopathogènes est celui des Entomophthoramycota. Ils présentent des cellules sporulées (conidiophores) spécialisées reposant sur une croissance de type phototropique positive (en présence de lumière). Les spores sont généralement évacuées de manière active par plusieurs mécanismes différents. Ces pathogènes forment fréquemment des épizooties (maladie frappant l’ensemble d’une population d’une espèce donnée dans une région plus ou moins vaste) tuant un grand nombre d’insectes dans de petites parcelles de forêts ou de systèmes agricoles.
4.2. Espèces entomopathogènes du groupe des Entomophthoramycota
Le taxon des Entomophthoramycota est composé principalement d’agents pathogènes d’insectes, peu infectent d’autres invertébrés, d’algues desmides et de gamétophytes de fougères, certains développent au contraire un mode de vie saprophytes.
Les espèces entomopathogènes sont réparties entre 19 genres : Entomophthora, Conidiobolus, Entomophaga, Erynia, Meristacrum, Neozygites, Strongwellsea, Massospora, Pandora, Eryniopsis (figure 4 ci-dessous), Batkoa, Tarichium, Completoria, Ballocephala, Zygnemomyces, Ancylistora, Ancylistes et Basidiobulus.
Il est difficile d’estimer le nombre d’entomopathogènes existants car nombre de ces genres infectent plusieurs hôtes, et leur taxonomie est en constante évolution. Cependant, une forte diversité morphologique et de stratégies a été décrite au sein de ce groupe.
Les champignons entomophthoroïdes attaquent de manière générale les insectes adultes, d’autres comme E. aquatica, E. conglomerata et Erynia aquatica infectent des larves de moustiques.
La transmission s’effectue par des spores diffusées dans l’environnement de manière active à l’exception d’un seul genre (Massospora) qui libère passivement ces spores alors que l’hôte demeure en vie. D’autres genres comme Strongwellsea et certaines espèces d’Entomophthora, d’Erynia et d’Entomophaga produisent des spores à l’intérieur ou l’extérieur des hôtes avant leur mort. Ces champignons, biotrophiques, ne développent pas de croissance somatique après la mort de l’insecte.
Le cas de Strongwellsea castrans infectant des Diptères Hylemya brassicae et H. platura (Anthomyiidae) sont des exemples classiques de ces situations où la sporulation se produit alors que l’hôte est encore en vie. Les insectes infectés sont caractérisés par la présence d’un large trou circulaire sur le côté latéral de l’abdomen.
C’est l’une des différences majeures avec les champignons hémibiotrophes (Ascomycètes) passant par une phase de parasitisme (biotrophique) et une phase saprophyte à l’intérieur ou à l’extérieur de l’hôte.
Chose surprenante, les insectes infectés peuvent être observés agissant normalement malgré le grand trou dans leur corps et rempli de tissu fongique et de conidiophores (cellules productrices de spores). Les mâles et les femelles ont été décrits comme infectés par S. castrans provoquant la castration et la mort prématurée.
Un autre cas similaire se produit avec Massospora cicadina qui se rencontre chez les cigales (Hémiptères) (figure ci-dessous). Ce champignon déclenche également la sporulation lorsque l’hôte est vivant, engendrant parfois l’effondrement de l’abdomen sous le poids du tissu fongique, voire sa désagrégation totale, ne restant in fine que la tête et le thorax de l’insecte. L’insecte conserve toutefois sa capacité de voler, augmentant ainsi la dispersion des spores dans l’environnement.
Bien que le groupe Mucoromycotina soit le plus grand et le plus diversifié au niveau morphologique des Zygomycètes, une seule espèce est entomopathogène : S. umbellata retrouvé sur des spécimens du genre Umbonia (Hemiptères) en Equateur et des papillons du genre Acraea (Nymphalidae) à Taïwan.
Figure 4 : A gauche -Cycle de développement de Eryniopsis lampyridarum – A) le coléoptère cantharide infecté par E. lampyridarum est mort avec les mandibules attachées à une tiges ou à une fleur, les élytres et les ailes s’ouvrent progressivement à mesure que le champignon se développe dans le corps de l’hôte – B) les conidiophores émergent directement du corps de l’hôte – C) conidium primaire – D) conidium primaire portant un conidium secondaire mature à l’extrémité – E) conidie secondaire – F) les conidies secondaires produiront éventuellement des capilliconidies en l’absence d’un hôte approprié – G) un autre coléoptère cantharide sera infecté s’il touche l’hymémium fongique exposé
A droite -Cycle de développement de Massospora cicadina – A) dispersion de spores par une cigale vivante infectée dont le corps se désintègre par l’activité fongique – B) cellules productrices de spores (conidiophores) à différents stades de développement – C) mouche présentant un trou sur l’abdomen causé par l’infection fongique – D) conidiophores présentant une spore terminale (Source : Araujo & Hughes, 2016)
5. Basidiomycètes
5.1. Généralités
Ce groupe contient des espèces connues pour les maladies qu’ils provoquent sur des végétaux cultivés : maladie du « charbon », ou de la « rouille ». Les Basidiomycètes se caractérisent par la formation de spores sexuelles (basidiospores) se formant à l’extérieur des cellules reproductrices spécialisées appelées basides. Ces spores sont dans la plupart des cas rejetées de force par des structures spécialisées.
Les Basiodiomycètes présentent certains traits écologiques importants. Ils colonisent le bois mort, décomposant la cellulose et la lignine, agissant également comme des décomposeurs de litière de feuilles sur le sol de la forêt. Les espèces pathogènes des végétaux provoquent d’importants dégâts dans les cultures, mais aussi en environnement forestiers : Armillariella mellea infectant de nombreuses espèces d’arbres et Heterobasidion annosum s’attaquant spécifiquement aux confères. Les espèces pathogènes des animaux comme certaines du genre Nematoctonus sont connues pour parasiter les nématodes.
5.2. Espèces entomopathogènes
Bien que ce phylum soit composé de 31 000 espèces décrites réparties en 1 500 genres, seuls trois d’entre eux sont connus pour infecter les insectes : Fibularhizoctonia infectant les œufs de termites, Uredinella (U. coccidiophaga et U. spinulosa) et Septobasidium (240 espèces) infectant des cochenilles (Hemiptères).
L’ordre des Septobasidiales (Uredinella et Septobasidium) présente une relation particulière et complexe avec leurs hôtes : les Diaspididae (Hemiptera).
Photo 3 : Cochenilles diaspines recouvertes par un champignon Septobasidium sp. (Source : Taylor Williams – NCCE)
Les Diaspididae (cochenilles diaspines – Coccoidea) sont de petits insectes phytophages sédentaires qui passent leur vie entière au même endroit sur une plante, conséquence de leur structure buccale de type suceuse. Pour se protéger, ne pouvant ni se déplacer et ne pouvant survivre sans protection, les juvéniles commencent à sécréter de fins fils de cire blanche qui, dans les 24 premières heures après l’éclosion, formeront une couche protectrice autour de l’insecte. Cependant, les larves demeurent exposées aux facteurs environnementaux.
Toutefois, des champignons du genre Septobasidium, pouvant créer des structures de plus de 20cm composées de tunnels et de chambres, offrent une structure défensive à vie pour les insectes (Figure 5) (photo 3). En contrepartie, le champignon infecte certains individus de la colonie de Diaspididae, provoquant leur nanisme ou la castration. L’atrophie des larves résulte d’une sous-alimentation provoquée par le détournement des nutriments provenant de la sève de la plante hôte par le champignon. Les adultes, également maintenus au réseau fongique par des hyphes, ne peuvent pas quitter le système.
Ce cas fourni un exemple de champignon cultivant des insectes !
Les deux espèces du genre Uredinella (U. spinulosa et U. coccidiophaga) peuvent être différenciées par la forme de leurs spores et du substrat dans lequel elles infectent leurs hôtes : feuilles et tronc pour U. spinulosa et uniquement le tronc pour U. coccidiophaga. Contrairement à Septobasidium qui présente une durée de vie indéfinie, puisque son corps est « renouvelé » à chaque saison par l’infection de nouveaux individus, le genre Uredinella infecte quant à lui uniquement le corps d’un seul insecte, la mort de l’insecte est donc associée à celle du champignon. Les spores sont produites au printemps et atteignent 0,2 à 1,5mm de diamètre.
Un autre cas de parasitisme de Basidiomycètes est celui entre certaines espèces du genre Fibularhizoctonia et des termites souterrains Reticulitermes. Il a été retrouvé parmi les tas d’œufs de ces termites des structures globuleuses (sclérotes) ayant le même diamètre et la même texture que les œufs, imitation telle que les ouvriers s’en occupent comme de leurs propres œufs. Il est suggéré que le champignon soit entretenu par les termites, et qu’il devienne pathogène dès lors que les termites cessent de s’en occuper.
Figure 5 : Cycle de développement de Septobasidium sp. (Source : Wikimedia.org)
6. Ascomycètes
6.1. Généralités
Le phylum des Ascomycètes est le plus grand groupe du règne des champignons avec plus de 64 000 espèces décrites. La plupart d’entre elles sont filamenteuses, produisant régulièrement des hyphes cloisonnés. Ils se caractérisent par la formation de spores sexuelles (ascospores) dans des structures en forme de sac appelés asques. Leur diversité écologique comprend des décomposeurs, des agents pathogènes de plantes, d’humains et d’animaux, ainsi que des relations mutualistes (lichens par exemple).
La majorité des espèces entomopathogènes possède une phase parasitaire bien développée infectant le corps de l’hôte. De plus, après avoir tué l’insecte, ce groupe est capable de coloniser le cadavre en passant à la nutrition saprophytique (hémibiotrophique), tout en maintenant la croissance de l’hyphe. Ces entomopathogènes seraient apparus et diversifiés dans les premières forêts tropicales humides.
La grande majorité des ascomycètes entomopathogènes forment leurs spores à l’intérieur de structures appelées péithèces.
6.2. Espèces entomopathogènes
Ce groupe comprend de nombreux champignons entomopathogènes (Pleosporales, Myriangiales, Ascosphaerales et ceux plus connus Hypocreales). Pour chacun des taxons, l’insecte meurt avant que le champignon débute sa phase de reproduction.
Ainsi, au sein des Pleosporales, les espèces entomopathogènes appartiennent au genre Podonectria, partageant des aspects inhabituels comme une coloration brillante, infectent des cochenilles, couvrant l’ensemble du corps d’une croûte de coton sur laquelle des spores multiseptées (cloisonnés) sont produits.
Les Myriangiales comprennent un certain nombre d’espèces associées aux plantes et aux cochenilles. Les espèces entomopathogènes présentent une croissance pérenne durant plusieurs années ou au moins jusqu’à la mort de la branche infestée par manque de nourriture. Les insectes hôtes sont remplis et recouverts de mycélium, la reproduction du champignon repose sur la production d’ascospores en absence de conidies (spores asexuées).
Le genre Ascosphaera (30 espèces) constitue un groupe d’agents pathogènes associés aux abeilles. La plupart des espèces sont saprophytes, se nourrissant exclusivement de miel, de cocons, des excréments larvaires ou les matériaux de nids tels que les feuilles, la boue ou la cire d’abeilles. Cependant, certaines espèces sont connues comme agents fongiques attaquant le couvain de nombreuses espèces d’abeilles solitaires et sociales, provoquant une maladie appelée « couvée de craie ».
Figure 6 : Cycle de développement de Ophiocordyceps unilateralis – A) les fourmis quittent le nid pour se nourrir sur le sol de la forêt – B) infections par des ascospores d’O. unilateralis – C et D) la fourmi quitte le nid environ 10 jours après la contamination pour aller mourir à une feuille ou une tige. Le champignon place la fourmi à un endroit précis optimal pour la dispersion des spores – E) 2 à 8 semaines (selon les conditions environnementales) après la mort de la fourmi, le champignon commence à produire des spores et à les disperser – F) 24h à 72h après être tombés sur le sol, les spores germent et forment des spores secondaires, les capilliconidiospores (Source : Araujo & Hughes, 2016)
L’infection survient lorsque la larve ingère des spores fongiques, le champignon se développe ensuite sous forme d’hyphes dans le corps de l’insecte, la tue et diffuse des spores à la surface de la cuticule.
La morphologie des Ascosphaera est particulièrement singulière par rapport à d’autres groupes fongiques. Les ascospores sont contenues dans une seule cellule agrandie formant un kyste brun à brun noirâtre : l’ascome. Leurs spores présentent également une curieuse similitude avec les grains de pollens.
Photo 4 : Fourmi « zombie » morte par infestation d’un champignon Ophiocordyceps sp. – Guyane française – Station des Nouragues (Source : B. Gilles – 2009)
Les Hypocreales englobent des genres importants d’entomopathogènes : Cordyceps, Tolypocladium, Hypocrella, Ophiocordyceps, Moelleriella, Samuelsia et Torrubiella.
Ces champignons sont connus pour infecter des mouches blanches (aleurodes – Aleyrodoidea) et les cochenilles des forêts tropicales, et provoquer des épizooties dans la population hôte. Peu de cas ont cependant été recensés en milieu subtropical.
Le genre Ophiocordyceps, en particulier les espèces attaquant les fourmis, est connu pour provoquer d’énormes infestations dans de petites zones appelées cimetières (figure 6 ci-dessus). En effet, l’un des phénomènes les plus fascinants concernant les champignons entomopathogènes est le comportement des fourmis zombies causé par Ophiocordyceps unilateralis. Décrite par Tulasne & Tulasne en 1865, l’espèce manipule de manière adaptative le comportement des fourmis ouvrières, obligeant les insectes à quitter la colonie pour trouver un site offrant un microclimat optimal pour que le champignon puisse produire ces ascospores. Les fourmis meurent en mordant fermement sur le dessous ou le bord d’une feuille, tige, branche, etc. (photo 4 ci-contre). La position que prend l’insecte à sa mort dépend de l’espèce de champignon (figure 7 ci-dessous).
Figure 7 : Photos de fourmis zombies – A) Camponotus atriceps infectée par Ophiocordyceps camponoti-atricipis – Amazonie brésilienne (Source : J.P.M. Araujo) – B) fourmi infestée par Desmidiospora myrmecophila (Source : M. Kukla) – C) Camponotus renggeri infestée par O. camponoti-renggeri (Source : J.P.M. Araujo) – D) Daceton armigerum infestée par O. dacetini (Source : J.P.M. Araujo) – E) fourmi infestée par Desmidiospora myrmecophila (Source : M. Kukla) – Source : Antwiki.org)
Tableau de synthèse des relations entre hôtes et entomopathogènes associés
Figure8 : Relations entre hôtes et entomopathogènes – à gauche : phylogénie des ordres d’insectes – à droite : phylogénie des groupes fongiques associés (Source : Araujo & Hughes, 2016)
Facteurs et mécanismes évolutifs à l’origine d’une diversité des entomopathogènes
La grande majorité des espèces d’entomopathogènes ont été recensées chez des Hemiptères suceurs de sèves. Par exemple, 180 espèces de champignons du genre Ascomycota infectent essentiellement des Hemiptères dont la plupart des cas sont des infections au stade adulte de l’insecte. Le groupe Hypocrellae Aschersonia (92 espèces) sont responsables de plus de la moitié des infections chez les Hemiptères des familles des Coccidae, des Aleyrodidae et des Lecaniidae.
Pourquoi les Hemiptères sont-ils des hôtes privilégiés pour les champignons entomopathogènes ?
Une étude de 2007 suggère que les Hypocreales auraient évolué à partir d’un ancêtre commun se développant au détriment de végétaux. Au cours de l’évolution, plusieurs transferts horizontaux (entre 5 et 8) auraient eu lieu entre la plante et l’insecte se nourrissant de la plante infectée.
Comment cela est-il survenu ?
Les insectes, apparus durant le Dévonien (les premiers fossiles datent de -407 millions d’années), ont connu deux événements majeurs de diversification. L’un, ayant permis l’apparition des principaux ordres d’insectes actuels, s’est produit avec le rayonnement des plantes à graines (spermaphytes) au Permien (-299-251 millions d’années), le second avec l’apparition des plantes à fleurs (angiospermes) au Crétacé inférieur (-130 millions d’années). Au cours de ce deuxième événement de diversification, des pièces buccales en forme de stylets longs et fins capables de créer une forte aspiration de la sève des plantes sont apparus chez un groupe d’insectes dont les descendants forment l’ordre des Hémiptères. Cette innovation a permis à ces insectes d’exploiter une nouvelle niche écologique sans concurrent.
Dans le même temps, certains champignons exploitant les tissus végétaux ont profité de cette proximité avec ces insectes pour effectuer un transfert vers ces derniers et profiter d’une nouvelle niche écologique (-190-150 millions d’années). La contamination avec divers insectes Hemiptères se serait effectuée par contact de proximité sur les végétaux hôtes. En effet, le groupe Hypocrellae Aschersonia présente des spores mitotiques adaptées à la dispersion à courte distance par la pluie sur la surface des feuilles.
Par la suite, le transfert se serait produit entre des insectes phytophages et des espèces qui ne l’étaient pas. Ces changements d’écologie ont donné naissance à trois des familles entomopathogènes les plus importantes : Clavicipitaceae (Hypocrella, Moelleriella, Samuelsia et Metacordyceps), Cordycipitaceae (Cordyceps et Torrubiella) et Ophiocordycipitaceae (Ophiocordycepsly, Tolypocladium). Cette stratégie a sans aucun doute été efficace comme l’illustre la diversité en espèces, la répartition mondiale et l’abondance écologique de ces champignons.
Un autre cas intéressant de diversité inhabituelle d’infections fongiques d’insectes est celui des mouches (Diptères). Bien que le nombre d’espèces concernés soit plus faible que pour les Hemiptères, il est remarquable que les Diptères soient le seul ordre d’insectes présentant des cas d’infestations à tous les stades de développement : œufs, larves, pupes et adultes. Parmi les 6 grands groupes d’entomopathogènes fongiques, seul celui des Basidiomycètes n’enregistre aucune contamination de Diptères.
Pourquoi les mouches sont-elles des hôtes pour les champignons entomopathogènes ?
Une des raisons repose probablement sur le fait que les mouches soient l’ordre des insectes le plus diversifié sur le plan écologique : hématophages, endoparasites et ectoparasites de vertébrés et d’invertébrés, prédateurs de larves et d’adultes d’arthropodes, parasitoïdes, pollinisateurs, phytophages, saprophytes, etc. De plus, les larves se développent dans des environnement aquatiques, semi-aquatiques, terrestres (humus, bois, champignons, etc.). Le fait d’occuper une telle diversité de niches écologiques a exposé les mouches et à augmenter les risques d’infections par des pathogènes fongiques et Oomycètes évoluant dans ces milieux.
Il est cependant remarquable que certaines espèces de mouches puissent être contaminées par 5 ou 6 groupes d’entomopathogènes différents et à tous les stades de développement. Cependant, la diversité d’entomopathogènes s’attaquant aux Diptères demeure faible au vu de la diversité des mouches (plus de 150 000 espèces).
Au-delà des ordres d’insectes évoqués précédemment, les larves de Lépidoptères et de Coléoptères sont également sujettes à des infections par des champignons entomopathogènes (Ascomycètes). Les larves de ces insectes sont principalement infectées par près de 90 espèces des genres Cordyceps et Ophiocordyceps. Les adultes sont quant à eux peu touchés par ces pathogènes, seuls 10 et 11 mentions pour les Lépidoptères et les Coléoptères respectivement.
Quels facteurs favorisent une telle prédominance des infections aux stades larvaires ?
Un certain nombre de traits biologiques différents entre les larves et les adultes peuvent être cruciaux pour expliquer ce schéma : 1) niches écologiques différentes – En tant qu’holométaboles, les stades larvaire et adulte sont séparés sur le plan écologique, occupant des micro-environnements complétement différents, évitant ainsi la concurrence entre les juvéniles et les adultes, 2) prévisibilité dans les échelles de temps – les larves de Coléoptères et de Lépidoptères présentent généralement une mobilité modeste par rapport aux adultes, 3) taux d’alimentation – la larve a besoin de manger de grandes quantités de nourriture et de stocker cette nourriture afin de croître le plus rapidement possible, 4) protection (cuticule) – les larves possèdent une cuticule plus fine et perméable pour croître à un rythme élevé et cela serait impossible d’être contaminé si elles avaient l’exosquelette dur des Coléoptères adultes.
L’autre ordre holométabolique majeur infecté par les Hypocreales est celui des Hyménoptères (abeilles, fourmis et guêpes). La plupart des infections se font durant la phase adulte. La socialité chez un grand nombre d’espèces de cet ordre réduit les risques d’infections fongiques, notamment chez les larves, en raison d’un toilettage constant des larves et entre individus d’une même colonie.
Bibliographie
Araujo J.P.M. & Hughes D.P. (2016) : Diversity of entomopathogenic fungi : which groupes conquered the insect body ? Advances in Genetics, 94 (lien)
Passion Entomologie utilise des cookies sur son site internet et vous donne le contrôle sur ce que vous souhaitez activer.
Ce site internet utilise les cookies pour améliorer votre expérience utilisateur et à des fins statistiques et publicitaires. Les cookies nécessaires sont essentiels au bon fonctionnement du site internet. En les désactivant, cela peut nuire à votre expérience utilisateur.
Les cookies fonctionnels aident à exécuter certaines fonctionnalités telles que le partage du contenu du site Web sur les plateformes de médias sociaux, la collecte de commentaires et d'autres fonctionnalités tierces.
Les cookies de performance sont utilisés pour comprendre et analyser les indices de performance clés du site Web, ce qui contribue à offrir une meilleure expérience utilisateur aux visiteurs.
Cookie
Durée
Description
_gat
1 minute
Ce cookie est installé par Google Universal Analytics pour limiter le taux de demande et ainsi limiter la collecte de données sur les sites à fort trafic.
Les cookies analytiques sont utilisés pour comprendre comment les visiteurs interagissent avec le site Web. Ces cookies aident à fournir des informations sur le nombre de visiteurs, le taux de rebond, la source de trafic, etc.
Cookie
Durée
Description
CONSENT
2 ans
YouTube définit ce cookie via des vidéos YouTube intégrées et enregistre des données statistiques anonymes.
vuid
2 ans
Vimeo installe ce cookie pour collecter des informations de suivi en définissant un identifiant unique pour intégrer des vidéos sur le site Web.
_ga
2 ans
Le cookie _ga, installé par Google Analytics, calcule les données des visiteurs, des sessions et des campagnes et assure également le suivi de l'utilisation du site pour le rapport d'analyse du site. Le cookie stocke les informations de manière anonyme et attribue un numéro généré de manière aléatoire pour reconnaître les visiteurs uniques.
_gat_gtag_UA_48379736_2
1 minute
Défini par Google pour distinguer les utilisateurs.
_gid
1 jour
Installé par Google Analytics, le cookie _gid stocke des informations sur la façon dont les visiteurs utilisent un site Web, tout en créant un rapport d'analyse des performances du site Web. Certaines des données collectées incluent le nombre de visiteurs, leur source et les pages qu'ils visitent de manière anonyme.
Les cookies publicitaires sont utilisés pour fournir aux visiteurs des publicités et des campagnes marketing pertinentes. Ces cookies suivent les visiteurs sur les sites Web et collectent des informations pour fournir des publicités personnalisées.
Les cookies nécessaires sont absolument essentiels au bon fonctionnement du site Web. Ces cookies assurent les fonctionnalités de base et les fonctions de sécurité du site Web, de manière anonyme.
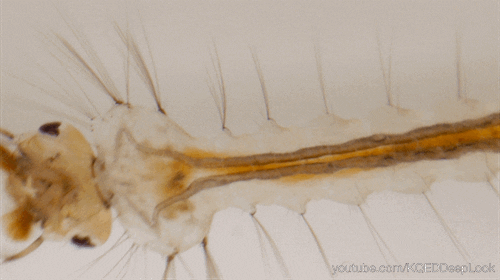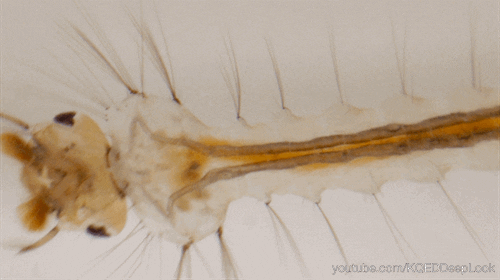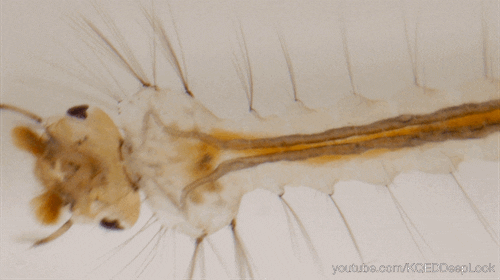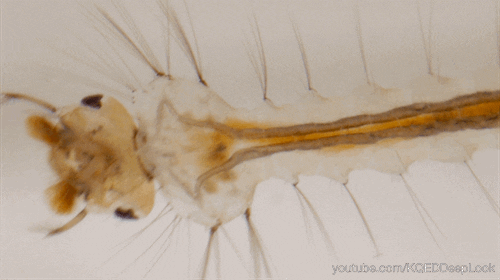
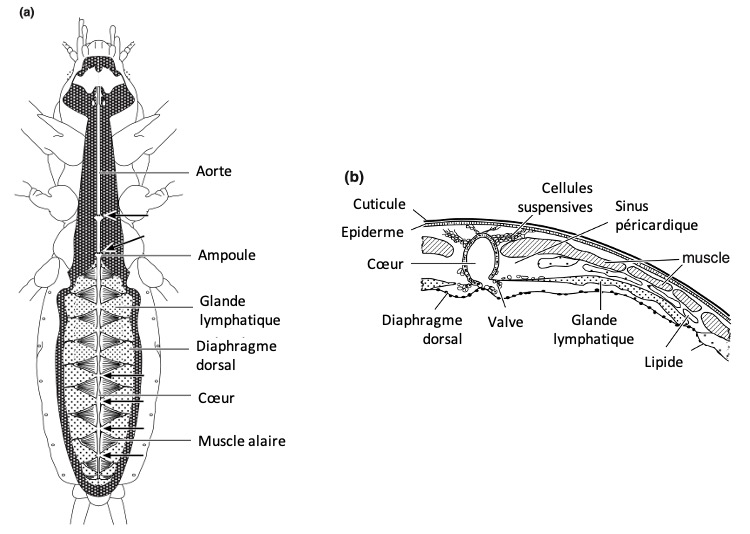
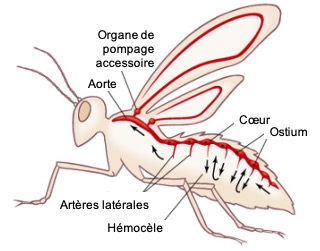
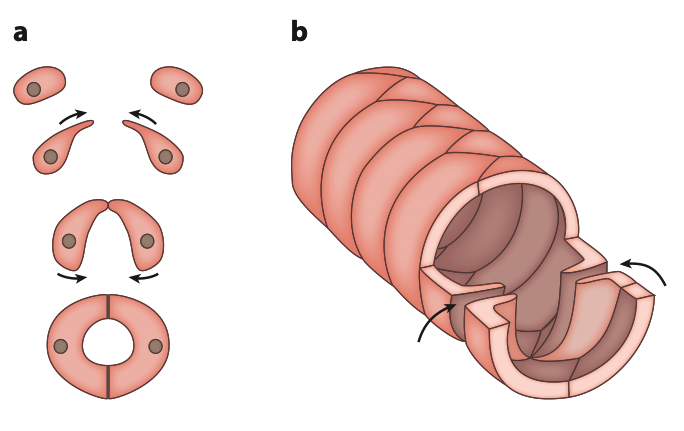
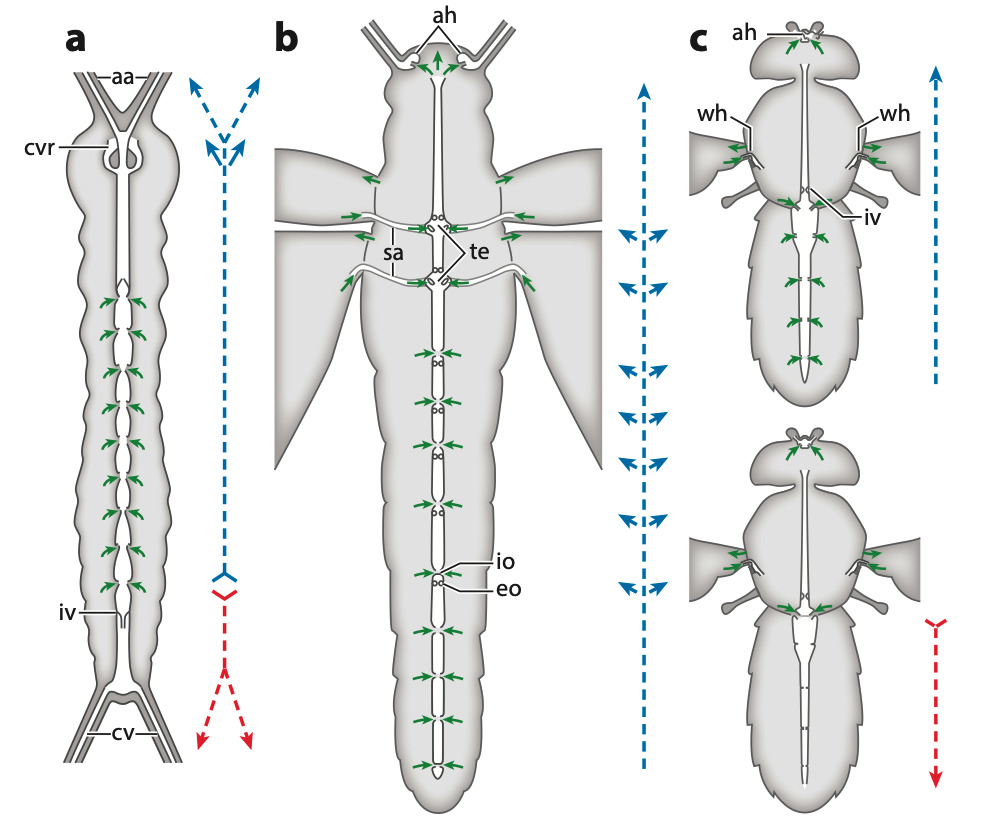
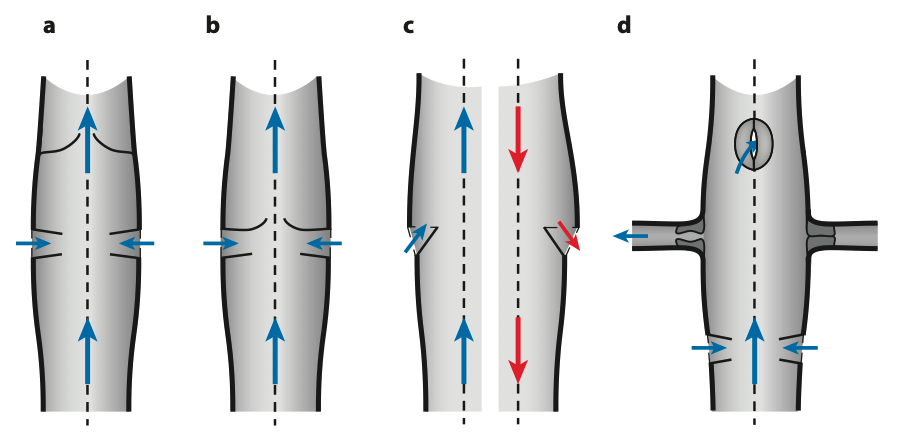
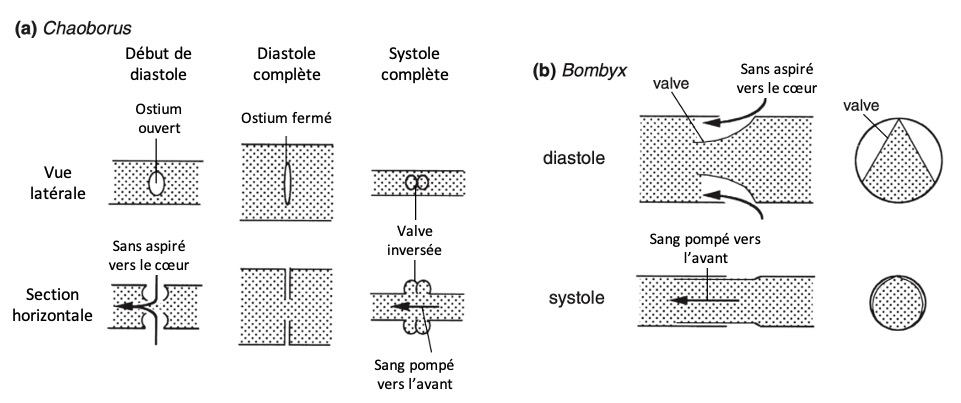
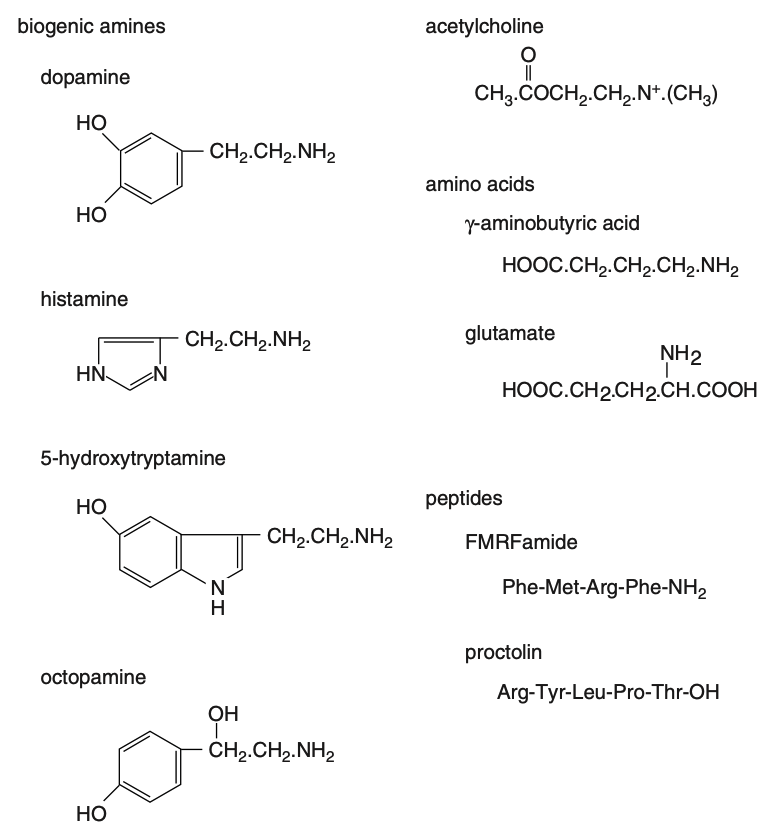
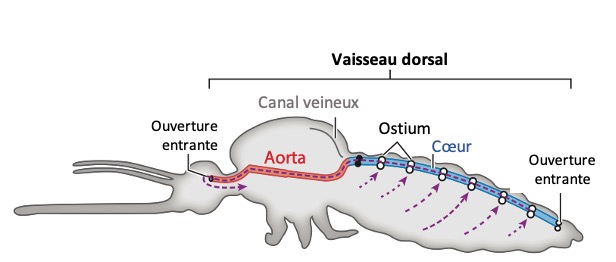
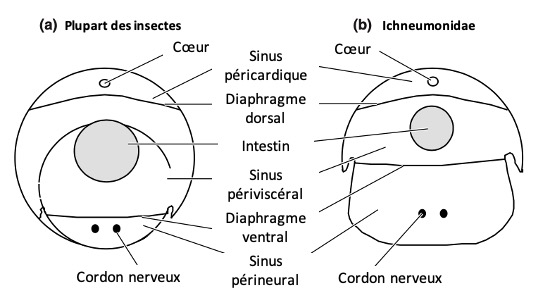
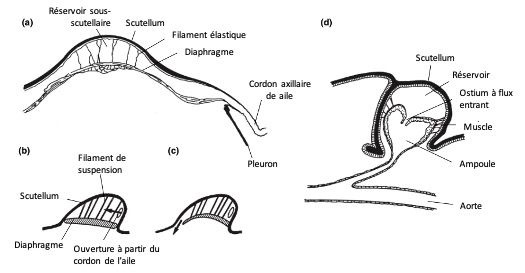
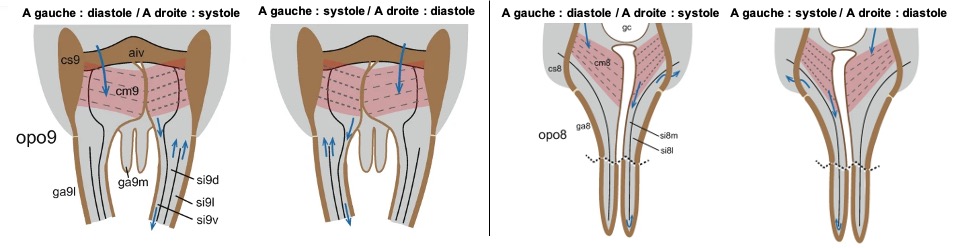
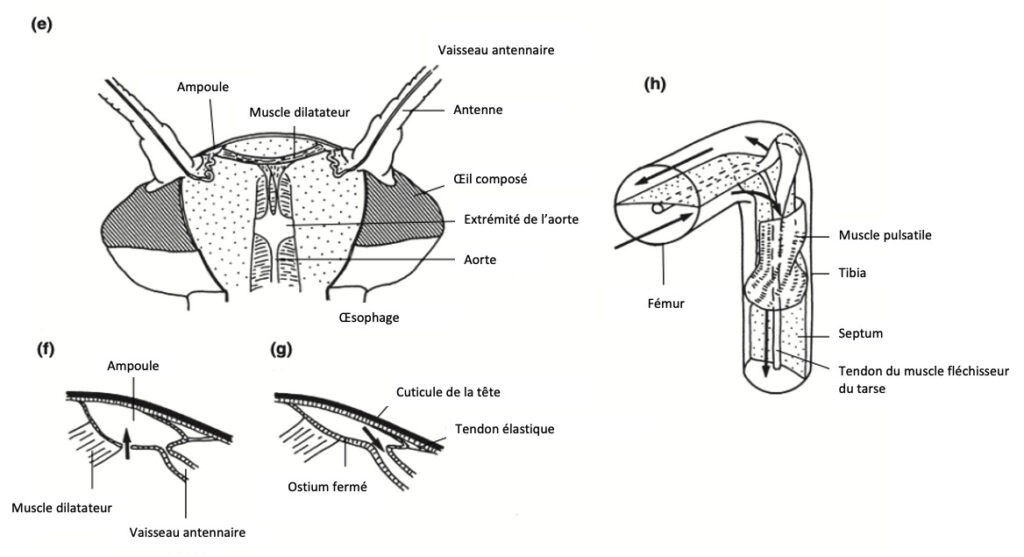
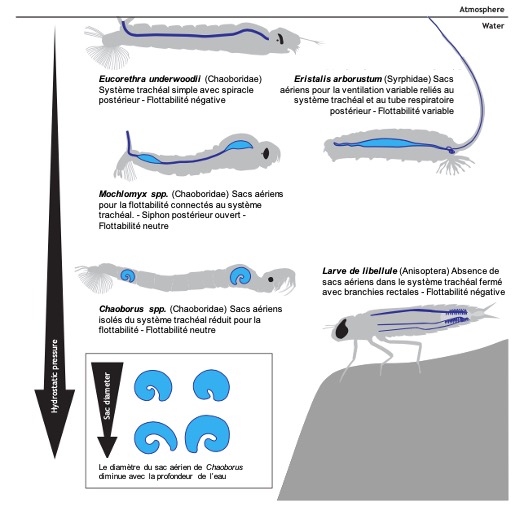
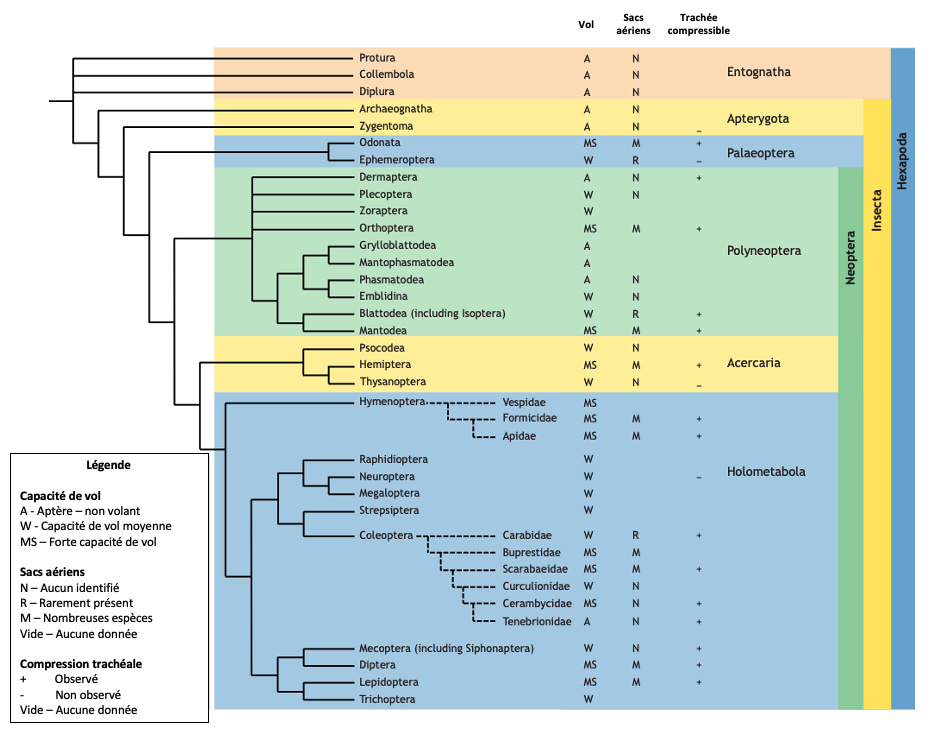
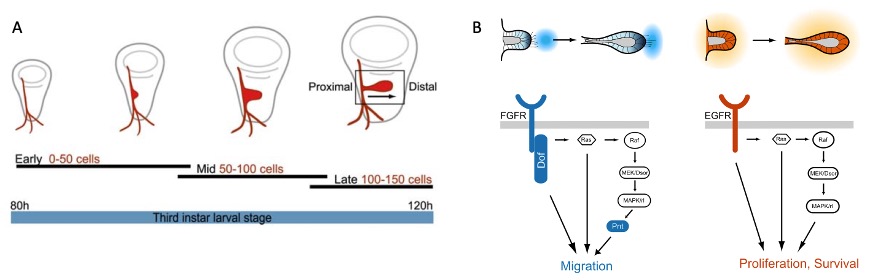
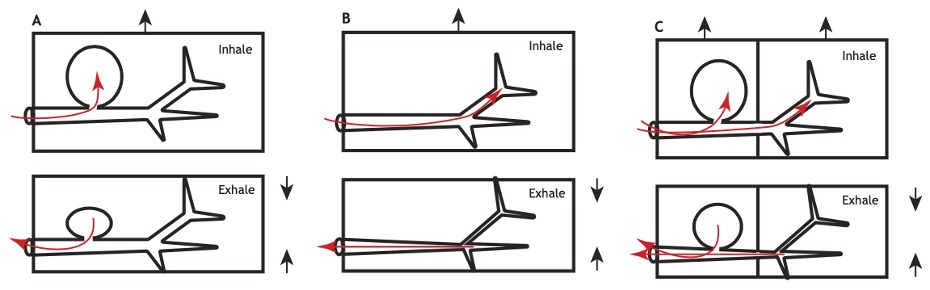
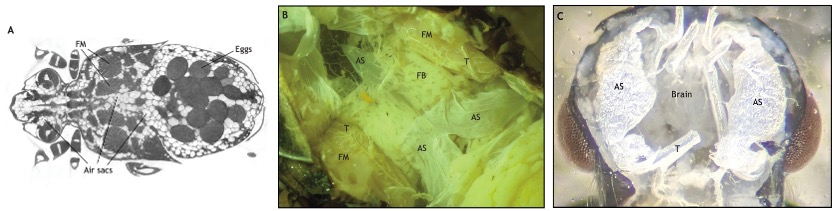
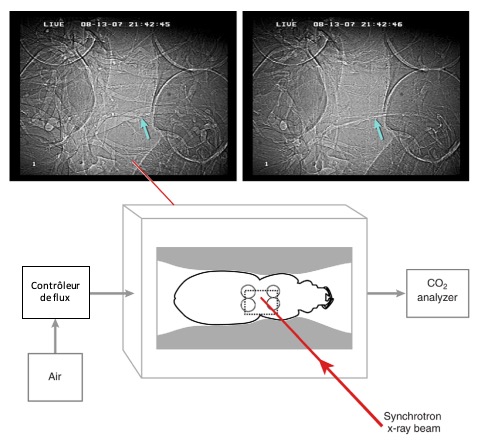
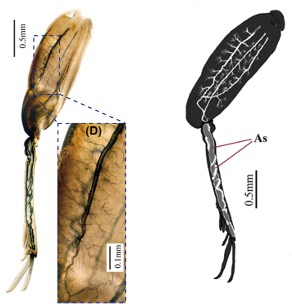
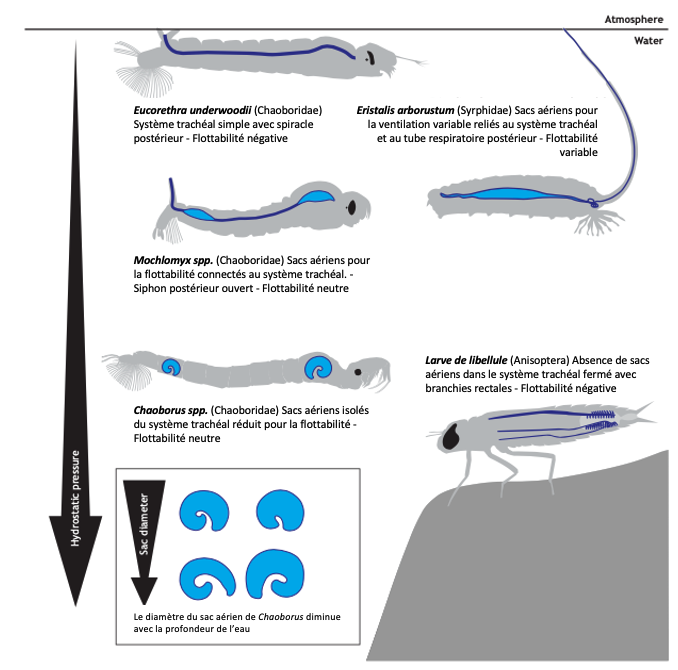

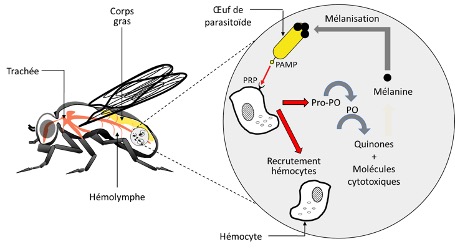
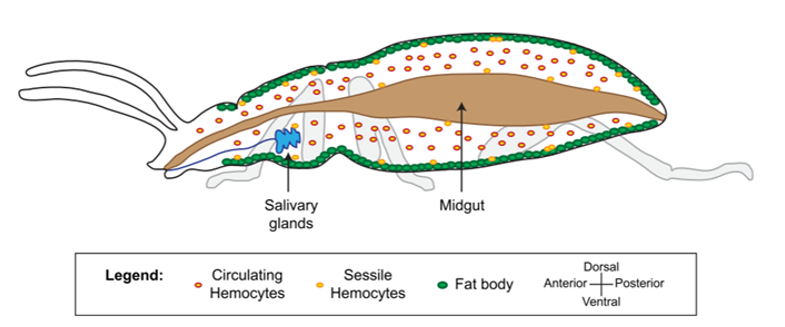
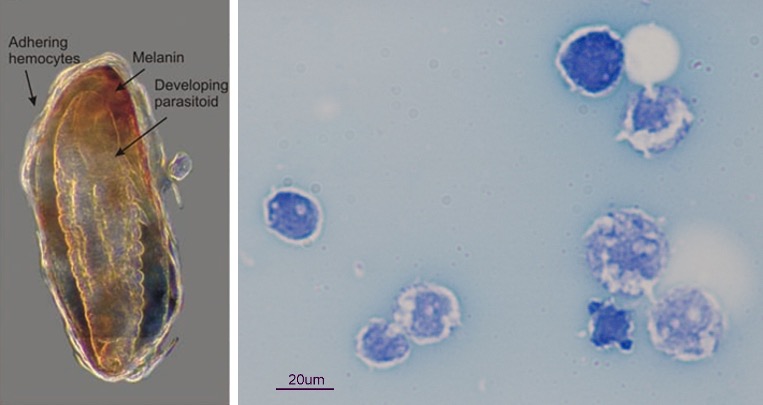

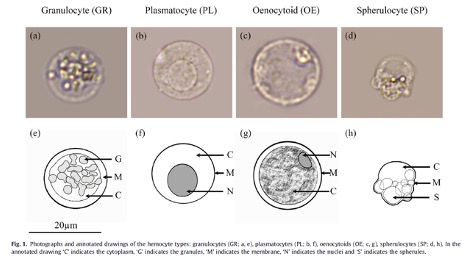
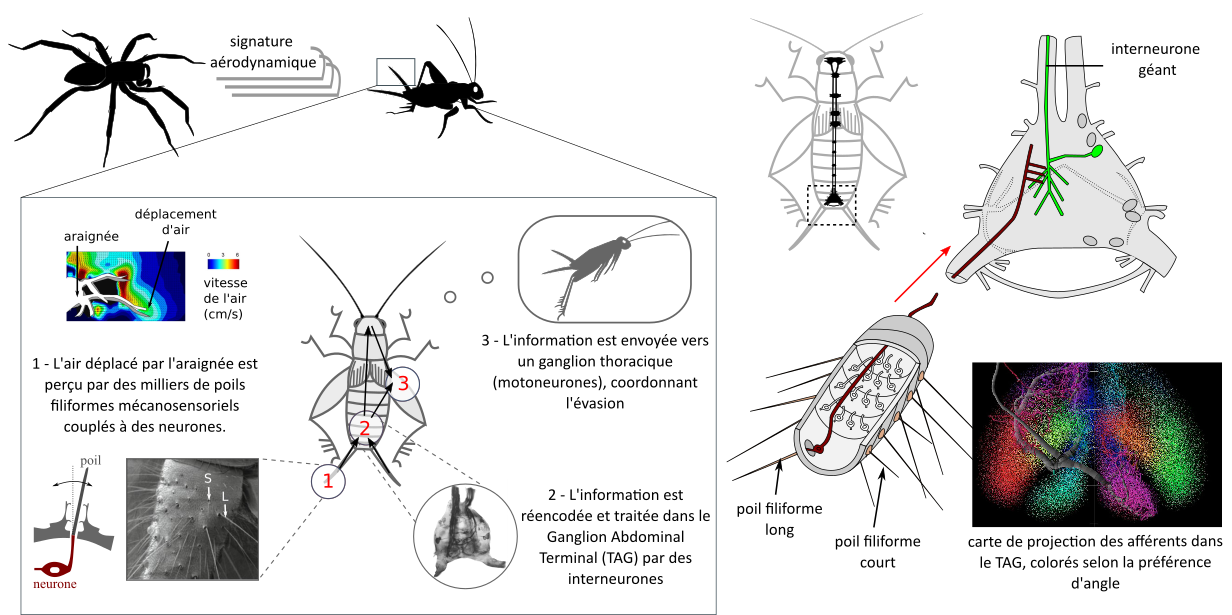
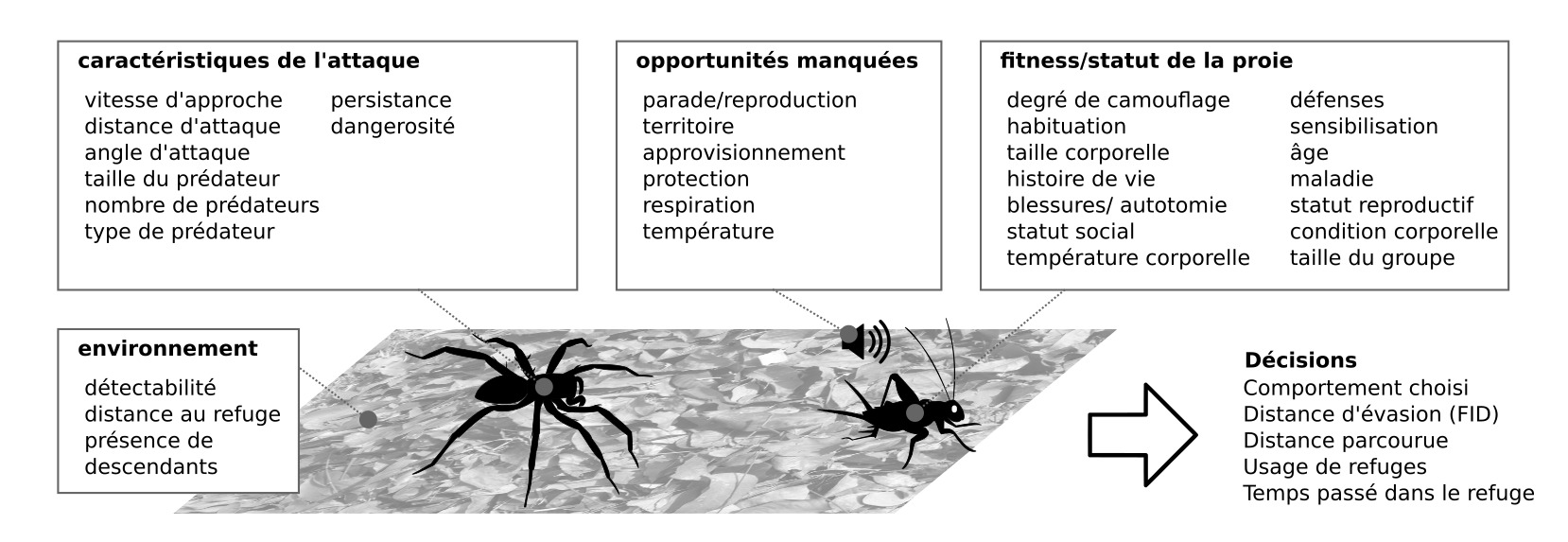
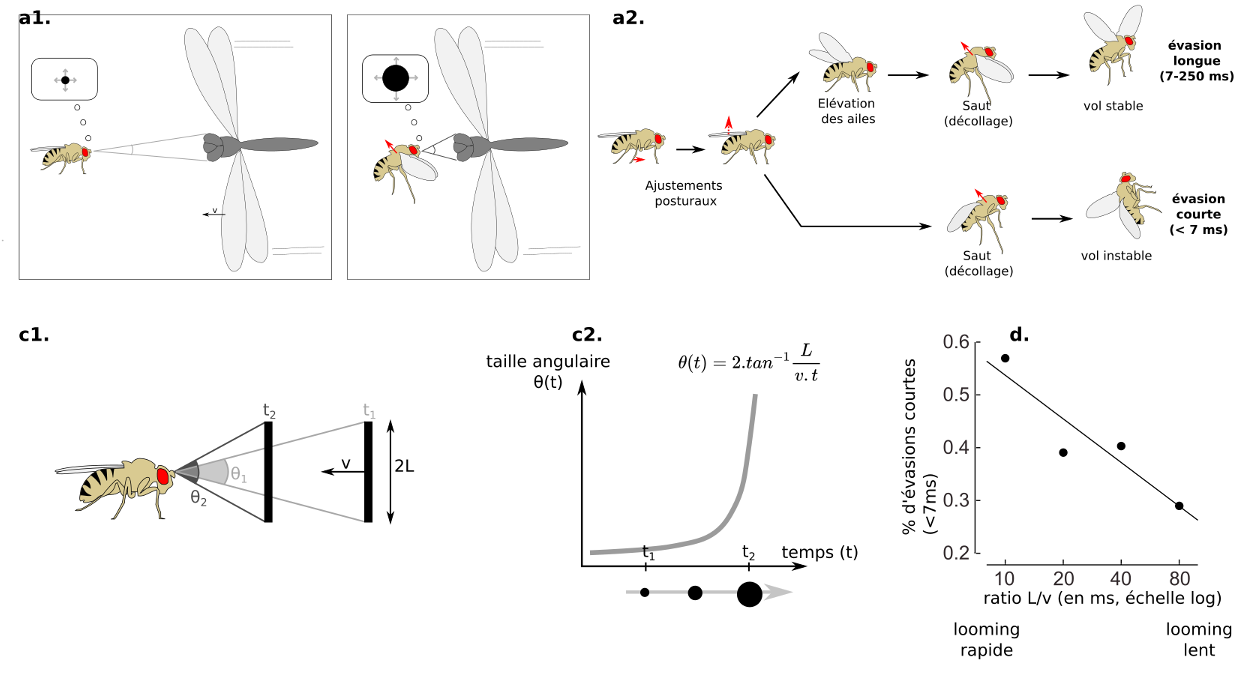
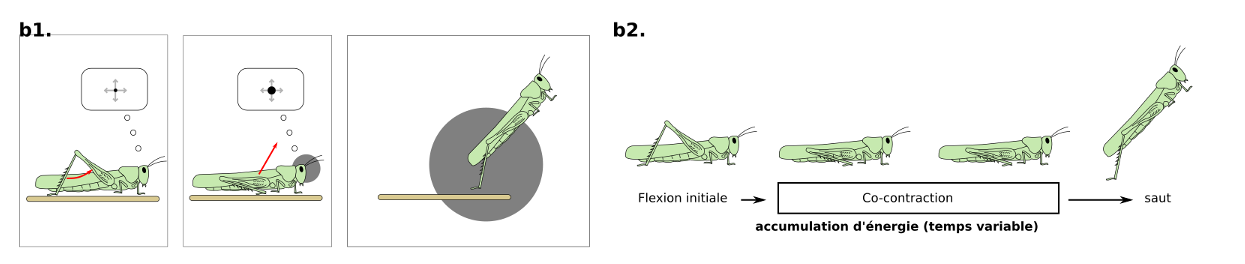
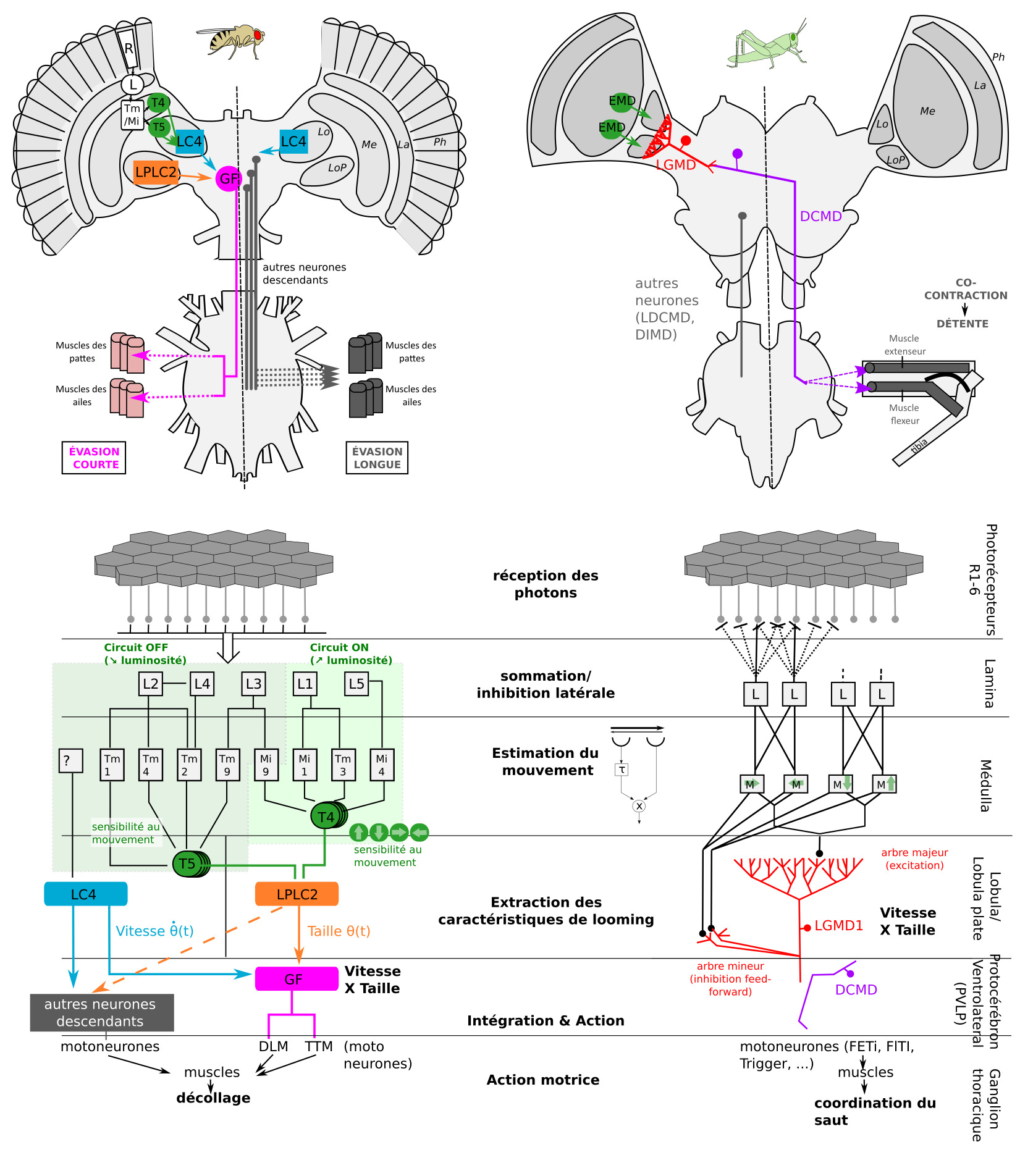

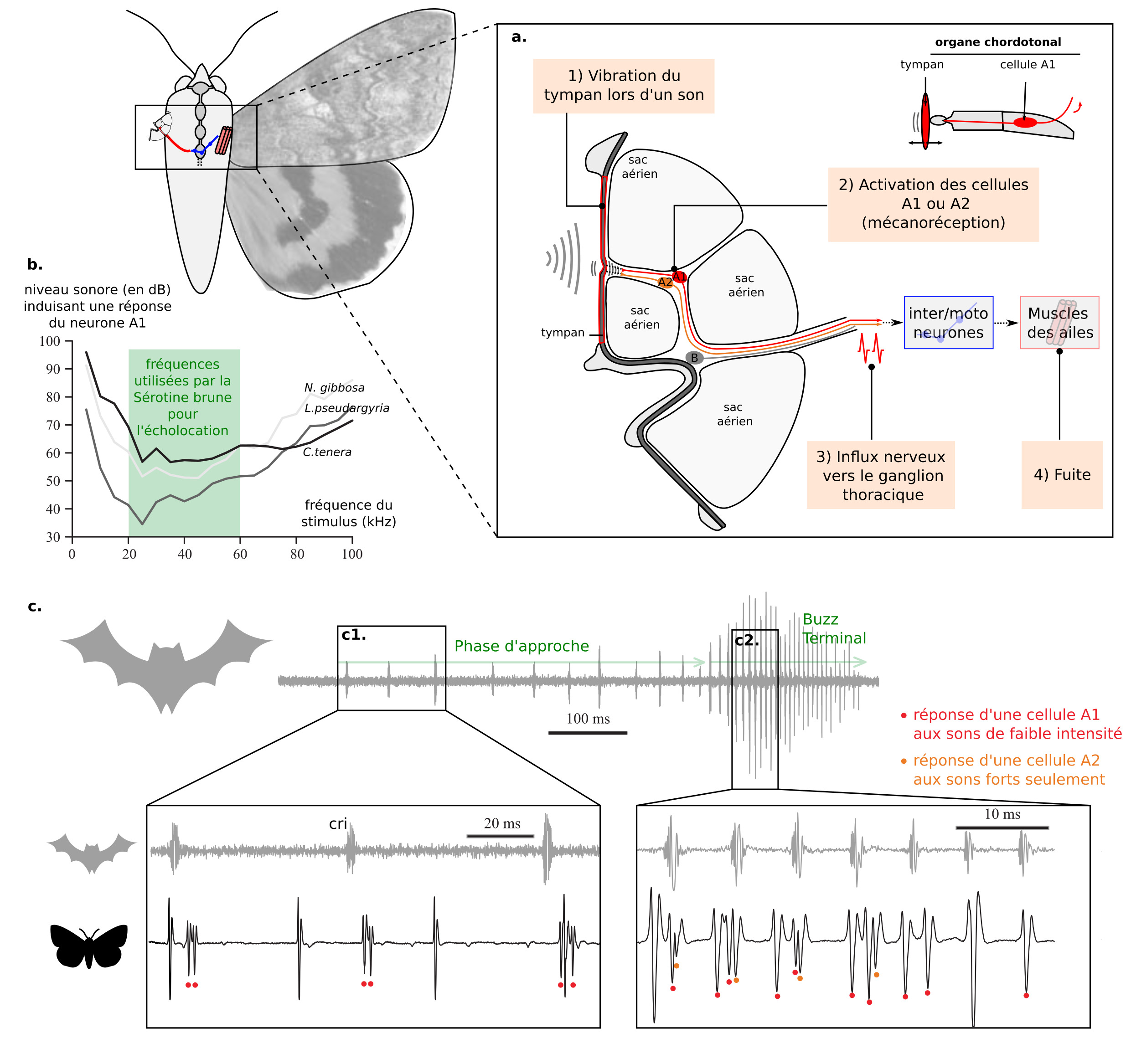
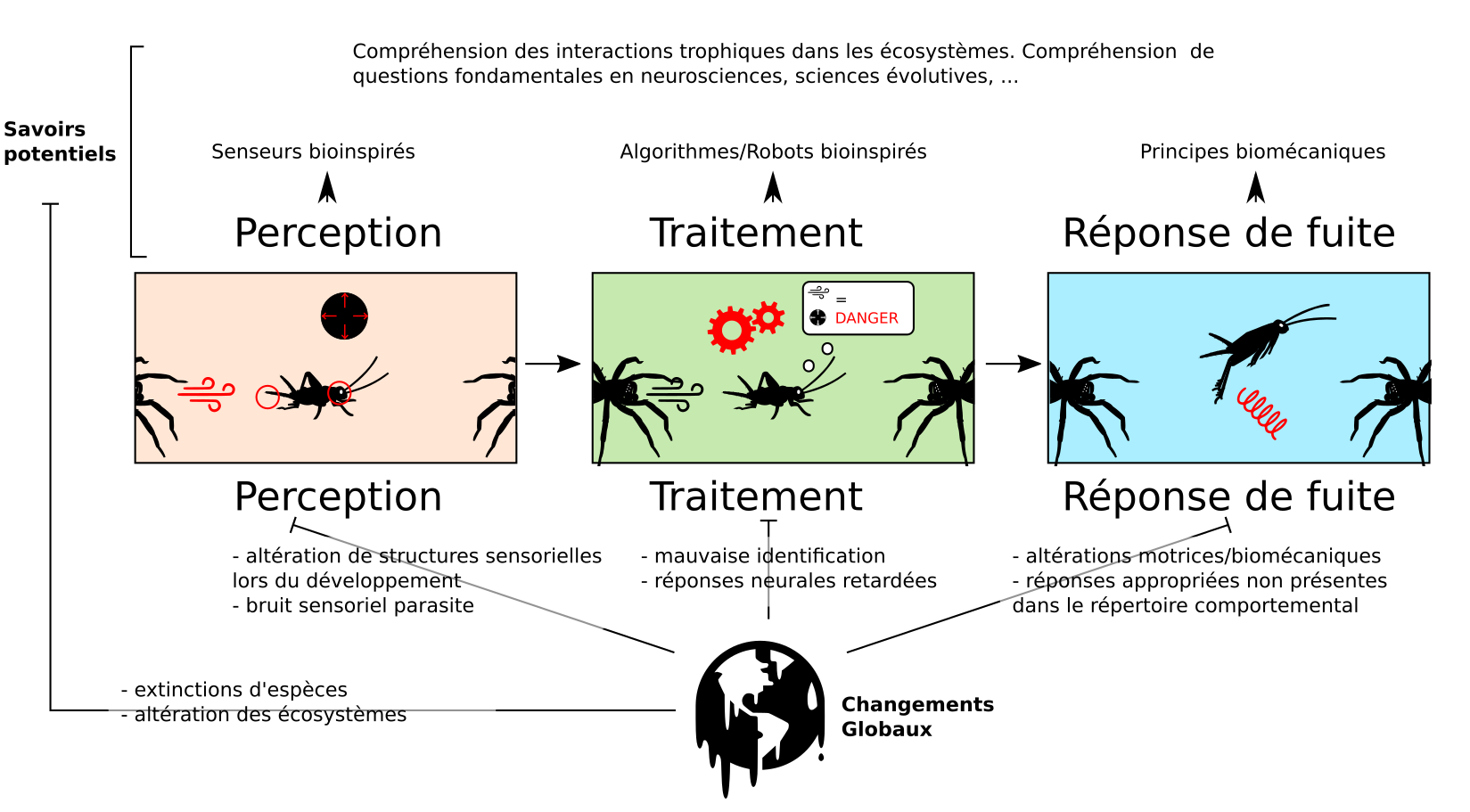
 Paul Clémençon
Paul Clémençon